Late Quaternary Environments, Denali National Park and Preserve, Alaska SCOTT A
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Joint Land Use Study
Fairbanks North Star Borough Joint Land Use Study United States Army, Fort Wainwright United States Air Force, Eielson Air Force Base Fairbanks North Star Borough, Planning Department July 2006 Produced by ASCG Incorporated of Alaska Fairbanks North Star Borough Joint Land Use Study Fairbanks Joint Land Use Study This study was prepared under contract with Fairbanks North Star Borough with financial support from the Office of Economic Adjustment, Department of Defense. The content reflects the views of Fairbanks North Star Borough and does not necessarily reflect the views of the Office of Economic Adjustment. Historical Hangar, Fort Wainwright Army Base Eielson Air Force Base i Fairbanks North Star Borough Joint Land Use Study Table of Contents 1.0 Study Purpose and Process................................................................................................. 1 1.1 Introduction....................................................................................................................1 1.2 Study Objectives ............................................................................................................ 2 1.3 Planning Area................................................................................................................. 2 1.4 Participating Stakeholders.............................................................................................. 4 1.5 Public Participation........................................................................................................ 5 1.6 Issue Identification........................................................................................................ -

Tanana Valley Stocked Lakes Fishing Guide
Tanana Valley Stocked Lakes Fishing Guide Alaska Department of Fish and Game Sport Fish Division April 2017 Table of Contents Tanana River Drainage Stocked Lakes Program .................................................................................................... 1 Fishing Facts and Tips ............................................................................................................................................ 3 Arctic Char (Salvelinus alpinus) ......................................................................................................................... 4 Arctic Grayling (Thymallus arcticus) ................................................................................................................. 5 Burbot (Lota lota) ............................................................................................................................................... 6 King (Chinook) Salmon (Oncorhynchus tshawytscha) and Silver (Coho) Salmon (Oncorhynchus kisutch) .... 7 Lake Trout (Salvelinus namaycush) ................................................................................................................... 8 Northern Pike (Esox lucius) ................................................................................................................................ 9 Rainbow Trout (Oncorhynchus mykiss) ........................................................................................................... 10 Road Guide .......................................................................................................................................................... -

National Geographic Trails Illustrated
Review (PDF) Denali National Park And Preserve (National Geographic Trails Illustrated Map) • Waterproof • Tear-Resistant • Topographic MapDenali National Park and Preserve covers a staggering six million acres of truly wild and pristine wilderness, bisected by a single road. The crown jewel of the park is the towering 20,320 foot Mount McKinley, North America’s tallest peak. Expertly researched and created in partnership with local land management agencies, National Geographic’s Trails Illustrated map of Denali National Park provides an unparalleled tool for exploring this remarkable region. Key areas of interest featured on this map include Denali State Park; Kroto, Moose Creek, and Lake Creek recreation rivers; Lake Minchumina; and the Kantishna Mining District. The print version of the map includes a detailed inset of the park entrance and headquarters.If you wish to camp in the park, a chart of campground information that includes the distance from the entrance, number and type of spaces, facilities, access, and contact information for making your reservations will help you plan your stay. Private vehicle travel in the park is limited, but this map’s coverage of the shuttle bus system will ensure you have the knowledge you need to access the entire park. Helpful information about regulations, avoiding close encounters with bears, hiking in the Kantishna Mining District, and safety tips, as well as clearly marked campgrounds, rental cabins, bush airstrips, trailheads, and more will help you enjoy all the park has to offer. The map base includes contour lines and elevations for summits, passes and many lakes.Every Trails Illustrated map is printed on "Backcountry Tough" waterproof, tear-resistant paper. -

Ground-Based Surveys for Dall's Sheep in Denali National Park
National Park Service U.S. Department of the Interior Natural Resource Stewardship and Science Ground-based Surveys for Dall’s Sheep in Denali National Park 2008-2017 Report Natural Resource Report NPS/DENA/NRR—2018/1611 ON THE COVER Observers search for Dall’s sheep during a ground survey on Mt. Margaret in 2016. NPS Photo Ground-based Surveys for Dall’s Sheep in Denali National Park 2008-2017 Report Natural Resource Report NPS/DENA/NRR—2018/1611 Bridget L. Borg1 1National Park Service Denali National Park and Preserve PO Box 9 Denali Park, AK 99755 March 2018 U.S. Department of the Interior National Park Service Natural Resource Stewardship and Science Fort Collins, Colorado The National Park Service, Natural Resource Stewardship and Science office in Fort Collins, Colorado, publishes a range of reports that address natural resource topics. These reports are of interest and applicability to a broad audience in the National Park Service and others in natural resource management, including scientists, conservation and environmental constituencies, and the public. The Natural Resource Report Series is used to disseminate comprehensive information and analysis about natural resources and related topics concerning lands managed by the National Park Service. The series supports the advancement of science, informed decision-making, and the achievement of the National Park Service mission. The series also provides a forum for presenting more lengthy results that may not be accepted by publications with page limitations. All manuscripts in the series receive the appropriate level of peer review to ensure that the information is scientifically credible, technically accurate, appropriately written for the intended audience, and designed and published in a professional manner. -

Regional Comprehensive Plan
FAIRBANKS NORTH STAR BOROUGH RREEGGIIOONNAALL CCOOMMPPRREEHHEENNSSIIVVEE PPLLAANN Adopted in September 2005 ~1947 LANCE MACKEY WINS THE 2005 YUKON QUEST A FREIGHT TEAM IN THE MID-1940S Fairbanks North Star Borough Regional Comprehensive Plan Adopted by the FNSB Borough Assembly September 13, 2005 (Ordinance No. 2005-56) Garry Hutchison, Presiding Officer Luke Hopkins Bonnie Williams Earl Romans Randy Frank Terry Aldridge Guy Sattley Hank Bartos Charlie Rex FNSB PLANNING COMMISSION MEMBERS Merle Jantz, Chair Chris Miller, Vice Chair Toby Hall Kelly Dryer Deborah Horner Gerald V. Neubert Alexandra M. Kienle Roy Earnest Steve Krause Bruce Hamilton FNSB Mayor Jim Whitaker FNSB Community Planning Department Bernardo Hernandez, Director 1 Community Planning Department P.O. Box 71267 Fairbanks, AK (907) 459-1260 www.co.fairbanks.ak.us Fairbanks North Star Borough Regional Comprehensive Plan Table of Contents Introduction Purpose 3 Vision 3 Process 3 Organization 4 History and Current Conditions 5 Plan Abstract 6 Plan Implementation Land Use 10 Regional Comprehensive Plan Map Definitions 15 Economic Development 18 Transportation and Infrastructure 21 Environment 24 Community and Human Resources 27 Appendix 31 Index 33 2 Fairbanks North Star Borough Regional Comprehensive Plan Introduction Purpose The Fairbanks North Star (FNSB) Regional Comprehensive Plan provides the foundation for future growth coupled with responsible stewardship of major attributes of the community in the perpetual evolving changes natural to man. The Plan provides the framework for citizens and officials to make decisions related to land use, and to form the basis for ordinances and programs to guide land development, and use. It is also a guide for responding to change in the community. -
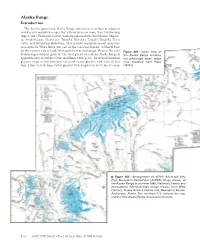
Alaska Range
Alaska Range Introduction The heavily glacierized Alaska Range consists of a number of adjacent and discrete mountain ranges that extend in an arc more than 750 km long (figs. 1, 381). From east to west, named ranges include the Nutzotin, Mentas- ta, Amphitheater, Clearwater, Tokosha, Kichatna, Teocalli, Tordrillo, Terra Cotta, and Revelation Mountains. This arcuate mountain massif spans the area from the White River, just east of the Canadian Border, to Merrill Pass on the western side of Cook Inlet southwest of Anchorage. Many of the indi- Figure 381.—Index map of vidual ranges support glaciers. The total glacier area of the Alaska Range is the Alaska Range showing 2 approximately 13,900 km (Post and Meier, 1980, p. 45). Its several thousand the glacierized areas. Index glaciers range in size from tiny unnamed cirque glaciers with areas of less map modified from Field than 1 km2 to very large valley glaciers with lengths up to 76 km (Denton (1975a). Figure 382.—Enlargement of NOAA Advanced Very High Resolution Radiometer (AVHRR) image mosaic of the Alaska Range in summer 1995. National Oceanic and Atmospheric Administration image mosaic from Mike Fleming, Alaska Science Center, U.S. Geological Survey, Anchorage, Alaska. The numbers 1–5 indicate the seg- ments of the Alaska Range discussed in the text. K406 SATELLITE IMAGE ATLAS OF GLACIERS OF THE WORLD and Field, 1975a, p. 575) and areas of greater than 500 km2. Alaska Range glaciers extend in elevation from above 6,000 m, near the summit of Mount McKinley, to slightly more than 100 m above sea level at Capps and Triumvi- rate Glaciers in the southwestern part of the range. -

Late Pleistocene /Early Holocene Site Structure In
LATE PLEISTOCENE /EARLY HOLOCENE SITE STRUCTURE IN BERINGIA : A CASE STUDY FROM THE BROKEN MAMMOTH SITE , INTERIOR ALASKA Kathryn E. Krasinski Department of Anthropology, University of Nevada, 1664 North Virginia Street MS 0096, Reno, NV 89557-0096; [email protected] David R. Yesner Department of Anthropology, University of Alaska Anchorage ABSTRACT Intrasite spatial analysis and the characterization of site structure at archaeological sites consider the relationship between archaeological material to derive connections between economic and social be- havior through the prehistoric use of space. This paper examines the spatial distribution of hearth fea- tures, débitage, and faunal remains in the late Pleistocene and early Holocene cultural components of the Broken Mammoth site, near Big Delta in interior Alaska. K-means analysis was used to reconstruct activity areas including lithic workshops, meal preparation areas, and refuse zones. Site organization suggests Broken Mammoth was a semipermanent base camp for early colonists of eastern Beringia. keywords: spatial analysis in archaeology, prehistoric activity areas, K-means cluster analysis, Beringia, Paleoindian archaeology, Broken Mammoth Site INTRODUCTION Most research on late Pleistocene and early Holocene sites merous hearth features in well-stratified contexts, relative in eastern Beringia has focused efforts on technological lack of postdepositional taphonomic disturbance, and fau- connections to Siberia and Northeast Asia (Goebel et nal preservation (Yesner et al. 1992). This paper addresses al. 1991; Hoffecker et al. 1993; West 1996; Yi and Clark the spatial structure of lithic and faunal patterns in the 1985). Studies of intrasite artifact distributions and site two oldest components of the Broken Mammoth site, cul- structure have seen much less research emphasis, but tural zones 3 and 4, dated between 12,000 and 9,300 14C a few notable exceptions exist (Higgs 1992; Hoffecker yrs bp. -

Tanana Lakes Recreation Area Master Plan Fairbanks North Star Borough Department of Parks & Recreation
Tanana Lakes Recreation Area Master Plan Fairbanks North Star Borough Department of Parks & Recreation Tanana Lakes Recreation Area Master Plan 2007 Planning Team: Fairbanks North Star Borough, Department of Parks & Recreation USKH, Inc. FNSB - Department of Parks and Recreation Executive Summary This Master Plan outlines the Fairbanks North Star Borough’s (FNSB’s) plan for the future development and use of the Tanana Lakes Recreation Area, a 750-acre multi-use park south of the City of Fairbanks along the Tanana River. The purpose of the Master Plan is to provide the FNSB with a long-term, planning guide for gravel extraction and development of the recreation area based on resource opportunities and constraints, development opportunities and constraints, and public needs. The Master Plan details the planning purpose and process, existing site conditions and land use, resources assessment, public process and demand, development plan, maintenance and operations considerations, and permits and authorizations required for development of the recreation area. The project concept originated approximately 15 years ago by the FNSB and its need to extract gravel for use at the South Cushman Landfill. The unsecured site has historically been a common place for illegal dumping of junk or abandoned vehicles and other refuse, as well as other unauthorized and criminal activities. Passing years have brought to the forefront the desire to clean up the site in order to enhance the wildlife habitat and natural features of the area in conjunction with the FNSB’s plan to extract gravel. As a result, this Master Plan has become a priority driven by both public safety and the importance of establishing this area for the community’s use and enjoyment. -

March 1St, 2021 Snow Water Equivalent
March 1, 2021 The USDA Natural Resources Conservation Service cooperates with the following organizations in snow survey work: Federal State of Alaska U.S. Depart of Agriculture - U.S. Forest Service Alaska Department of Fish and Game Chugach National Forest Alaska Department of Transportation and Tongass National Forest Public Facilities U.S. Department of Commerce Alaska Department of Natural Resources NOAA, Alaska Pacific RFC Division of Parks U.S. Department of Defense Division of Mining and Water U.S. Army Corps of Engineers Division of Forestry U.S. Department of Interior Alaska Energy Authority Bureau of Land Management Alaska Railroad U.S. Geological Survey Soil and Water Conservation Districts U. S. Fish and Wildlife Service Homer SWCD National Park Service Fairbanks SWCD Salcha-Delta SWCD Municipalities University of Alaska Anchorage Geophysical Institute Juneau Water and Environment Research Private Alaska Public Schools Alaska Electric, Light and Power, Juneau Mantanuska-Susitna Borough School Alyeska Resort, Inc. District Alyeska Pipeline Service Company Eagle School, Gateway School District Anchorage Municipal Light and Power Chugach Electric Association Canada Copper Valley Electric Association Ministry of the Environment Homer Electric Association British Columbia Ketchikan Public Utilities Department of the Environment Prince William Sound Science Center Government of the Yukon The U.S. Department of Agriculture (USDA) prohibits discrimination in all its programs and activities on the basis of race, color, nation- al origin, age, disability, and where applicable, sex, marital status, familial status, parental status, religion, sexual orientation, genetic information, political beliefs, reprisal, or because all or a part of an individual’s income is derived from any public assistance program. -

Denali National Park and Preserve
National Park Service U.S. Department of the Interior Natural Resource Program Center Water Resources Stewardship Report Denali National Park and Preserve Natural Resource Technical Report NPS/NRPC/WRD/NRTR—2007/051 ON THE COVER Photograph: Toklat River, Denali National Park and Preserve (Guy Adema, 2007) Water Resources Stewardship Report Denali National Park and Preserve Natural Resource Technical Report NPS/NRPC/WRD/NRTR-2007/051 Kenneth F. Karle, P.E. Hydraulic Mapping and Modeling P.O. Box 181 Denali Park, Alaska 99755 September 2007 U.S. Department of the Interior National Park Service Natural Resources Program Center Fort Collins, Colorado The Natural Resource Publication series addresses natural resource topics that are of interest and applicability to a broad readership in the National Park Service and to others in the management of natural resources, including the scientific community, the public, and the NPS conservation and environmental constituencies. Manuscripts are peer- reviewed to ensure that the information is scientifically credible, technically accurate, appropriately written for the intended audience, and is designed and published in a professional manner. The Natural Resources Technical Reports series is used to disseminate the peer-reviewed results of scientific studies in the physical, biological, and social sciences for both the advancement of science and the achievement of the National Park Service’s mission. The reports provide contributors with a forum for displaying comprehensive data that are often deleted from journals because of page limitations. Current examples of such reports include the results of research that addresses natural resource management issues; natural resource inventory and monitoring activities; resource assessment reports; scientific literature reviews; and peer reviewed proceedings of technical workshops, conferences, or symposia. -

Ground-Water Resources of the Fairbanks Area Alaska
Ground-Water Resources of the Fairbanks Area Alaska GEOLOGICAL SURVEY WATER-SUPPLY PAPER 1590 1863 Ground-Water Resources of the Fairbanks Area Alaska By D. J. CEDERSTROM GEOLOGICAL SURVEY WATER-SUPPLY PAPER 1590 Ground-water occurrence in an area of discontinuous permafrost UNITED STATES GOVERNMENT PRINTING OFFICE, WASHINGTON : 1963 UNITED STATES DEPARTMENT OF THE INTERIOR STEWART L. UDALL, Secretary GEOLOGICAL SURVEY Thomas B. Nolan, Director The U.S. Geological Survey Library has cataloged this publication as follows: Cederstrom, Dagfin John, 1908 Ground-water resources of the Fairbanks area, Alaska. Washington, U.S. Govt. Print. Off., 1962. iv, 84 p. illus., maps (part fold, in pocket) diagrs., tables. 24 cm. (U.S. Geological Survey. Water-supply paper 1590) Bibliography: p. 54-55. 1. Water-supply-Alaska-Fairbanks area. 2. Water, Underground- Alaska-Fairbanks area. 3. Water-Analysis. 4. Borings-Alaska-Fair banks area. (Series) For sale by the Superintendent of Documents, U.S. Government Printing Office Washington 25, D.C. CONTENTS Page Abstract_ _ _ _-_____--____-_______________________-_------__------ 1 Introduction. _____________________________________________________ 2 Scope and purpose of report____________________________________ 2 Geography___ _______________________________________________ 4 Topography ___________________________________________________ 4 Settlement and population________----_---_-___-_-_--_---------- 5 Climate ______________________________________________________ 6 Industry _____________________________________________________ -

Alaska Park Science Anchorage, Alaska
National Park Service U.S. Department of Interior Alaska Regional Office Alaska Park Science Anchorage, Alaska PROCEEDINGS OF THE CentrCentralal AlaskAlaskaa PParkark SciencSciencee SymposiumSymposium SeptemberSeptember 12-14,12-14, 2006 2006 Denali Park, Alaska Volume 6, Issue 2 Parks featured in this Table of Contents issue of Alaska Park Science Keynote Address Alaska Parks in a Warming Climate: Conserving a Changing Future __________________________ 6 S K A Yukon-Charley Rivers Synthesis L A National Preserve Crossing Boundaries in Changing Environment: Norton Sound A A Synthesis __________________________________________12 Monitoring a Changing Climate Denali National Park and Preserve Long-term Air Quality Monitoring Wrangell-St. Elias in Denali National Park and Preserve __________________18 National Park and Preserve Monitoring Seasonal and Long-term Climate Changes and Extremes in the Central Alaska Network__________ 22 Physical Environment and Sciences Glacier Monitoring in Denali National Park and Preserve ________________________________________26 Applications of the Soil-Ecological Survey of Denali National Park and Preserve__________________31 Bristol Bay Gulf of Alaska Using Radiocarbon to Detect Change in Ecosystem Carbon Cycling in Response to Permafrost Thawing____34 A Baseline Study of Permafrost in the Toklat Basin, Denali National Park and Preserve ____________________37 Dinosauria and Fossil Aves Footprints from the Lower Cantwell Formation (latest Cretaceous), Denali National Park and Preserve ____________________41