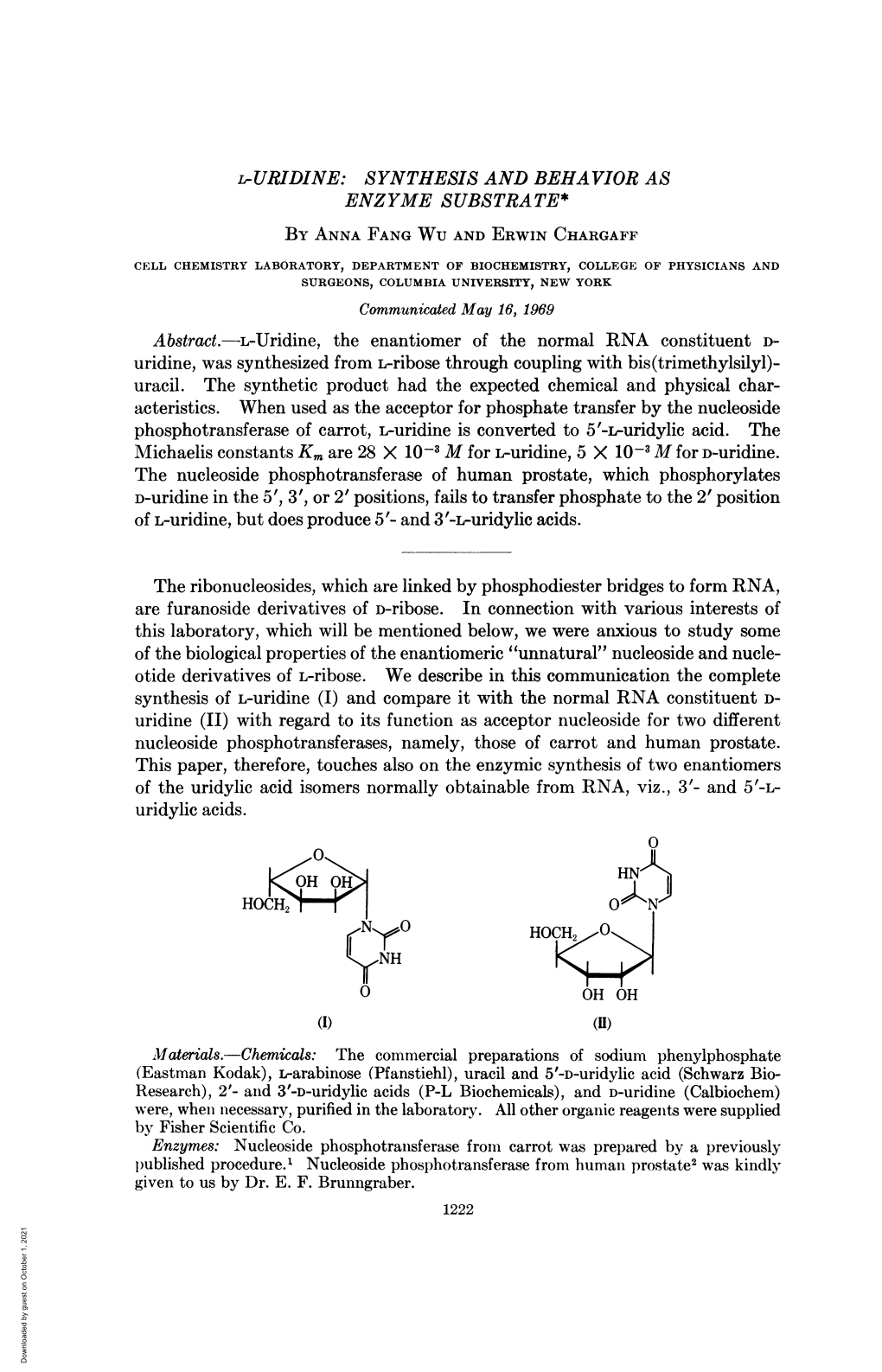
~~~~'0 HOCH, N~R0 OC NH~A HOC 3
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Nucleotide Degradation
Nucleotide Degradation Nucleotide Degradation The Digestion Pathway • Ingestion of food always includes nucleic acids. • As you know from BI 421, the low pH of the stomach does not affect the polymer. • In the duodenum, zymogens are converted to nucleases and the nucleotides are converted to nucleosides by non-specific phosphatases or nucleotidases. nucleases • Only the non-ionic nucleosides are taken & phospho- diesterases up in the villi of the small intestine. Duodenum Non-specific phosphatases • In the cell, the first step is the release of nucleosides) the ribose sugar, most effectively done by a non-specific nucleoside phosphorylase to give ribose 1-phosphate (Rib1P) and the free bases. • Most ingested nucleic acids are degraded to Rib1P, purines, and pyrimidines. 1 Nucleotide Degradation: Overview Fate of Nucleic Acids: Once broken down to the nitrogenous bases they are either: Nucleotides 1. Salvaged for recycling into new nucleic acids (most cells; from internal, Pi not ingested, nucleic Nucleosides acids). Purine Nucleoside Pi aD-Rib 1-P (or Rib) 2. Oxidized (primarily in the Phosphorylase & intestine and liver) by first aD-dRib 1-P (or dRib) converting to nucleosides, Bases then to –Uric Acid (purines) –Acetyl-CoA & Purine & Pyrimidine Oxidation succinyl-CoA Salvage Pathway (pyrimidines) The Salvage Pathways are in competition with the de novo biosynthetic pathways, and are both ANABOLISM Nucleotide Degradation Catabolism of Purines Nucleotides: Nucleosides: Bases: 1. Dephosphorylation (via 5’-nucleotidase) 2. Deamination and hydrolysis of ribose lead to production of xanthine. 3. Hypoxanthine and xanthine are then oxidized into uric acid by xanthine oxidase. Spiders and other arachnids lack xanthine oxidase. -

Chapter 23 Nucleic Acids
7-9/99 Neuman Chapter 23 Chapter 23 Nucleic Acids from Organic Chemistry by Robert C. Neuman, Jr. Professor of Chemistry, emeritus University of California, Riverside [email protected] <http://web.chem.ucsb.edu/~neuman/orgchembyneuman/> Chapter Outline of the Book ************************************************************************************** I. Foundations 1. Organic Molecules and Chemical Bonding 2. Alkanes and Cycloalkanes 3. Haloalkanes, Alcohols, Ethers, and Amines 4. Stereochemistry 5. Organic Spectrometry II. Reactions, Mechanisms, Multiple Bonds 6. Organic Reactions *(Not yet Posted) 7. Reactions of Haloalkanes, Alcohols, and Amines. Nucleophilic Substitution 8. Alkenes and Alkynes 9. Formation of Alkenes and Alkynes. Elimination Reactions 10. Alkenes and Alkynes. Addition Reactions 11. Free Radical Addition and Substitution Reactions III. Conjugation, Electronic Effects, Carbonyl Groups 12. Conjugated and Aromatic Molecules 13. Carbonyl Compounds. Ketones, Aldehydes, and Carboxylic Acids 14. Substituent Effects 15. Carbonyl Compounds. Esters, Amides, and Related Molecules IV. Carbonyl and Pericyclic Reactions and Mechanisms 16. Carbonyl Compounds. Addition and Substitution Reactions 17. Oxidation and Reduction Reactions 18. Reactions of Enolate Ions and Enols 19. Cyclization and Pericyclic Reactions *(Not yet Posted) V. Bioorganic Compounds 20. Carbohydrates 21. Lipids 22. Peptides, Proteins, and α−Amino Acids 23. Nucleic Acids ************************************************************************************** -

Differential Effects of the Poly (ADP-Ribose)Polymerase (PARP
British Journal of Cancer (2001) 84(1), 106–112 © 2001 Cancer Research Campaign doi: 10.1054/ bjoc.2000.1555, available online at http://www.idealibrary.com on http://www.bjcancer.com Differential effects of the poly (ADP-ribose) polymerase (PARP) inhibitor NU1025 on topoisomerase I and II inhibitor cytotoxicity in L1210 cells in vitro KJ Bowman*, DR Newell, AH Calvert and NJ Curtin Cancer Research Unit, University of Newcastle upon Tyne Medical School, Framlington Place, Newcastle upon Tyne NE2 4HH, UK Summary The potent novel poly(ADP-ribose) polymerase (PARP) inhibitor, NU1025, enhances the cytotoxicity of DNA-methylating agents and ionizing radiation by inhibiting DNA repair. We report here an investigation of the role of PARP in the cellular responses to inhibitors of topoisomerase I and II using NU1025. The cytotoxicity of the topoisomerase I inhibitor, camptothecin, was increased 2.6-fold in L1210 cells by co-incubation with NU1025. Camptothecin-induced DNA strand breaks were also increased 2.5-fold by NU1025 and exposure to camptothecin-activated PARP. In contrast, NU1025 did not increase the DNA strand breakage or cytotoxicity caused by the topoisomerase II inhibitor etoposide. Exposure to etoposide did not activate PARP even at concentrations that caused significant levels of apoptosis. Taken together, these data suggest that potentiation of camptothecin cytotoxicity by NU1025 is a direct result of increased DNA strand breakage, and that activation of PARP by camptothecin-induced DNA damage contributes to its repair and consequently cell survival. However, in L1210 cells at least, it would appear that PARP is not involved in the cellular response to etoposide-mediated DNA damage. -

Evidence Suggests That RNA Was a Product of Evolution
Putting together the pieces: Evidence suggests that RNA was a product of evolution Brian Cafferty and Nicholas V. Hud, Georgia Institute of Technology, Atlanta, GA, USA For the past four decades, prebiotic chemists have attempted to demonstrate the formation of RNA polymers by plausible prebiotic reactions. There have been notable advances, but to be certain, the spontaneous formation of RNA remains a grand challenge in origins of life research. From a different perspective, there are reasons to seriously consider the possibility that RNA is a product of evolution. If so, there may have never been a prebiotic mechanism that produced RNA polymers. We subscribe to this latter view and hypothesize that RNA is the penultimate member of continuous lineage of genetic polymers, with DNA being the ultimate member of this lineage. In this essay, we briefly summarize the case for why RNA is likely the descendant of one or more pre-RNA polymers that spontaneous assembled on the prebiotic earth. Nucleosides are each an assemblage of a nucleobase and a ribose sugar, whereas nucleotides, the monomeric units of RNA, are phosphorylated nucleosides (Figure 1). Prebiotic chemists have typically sought to form RNA in a seQuential fashion, starting with the formation of nucleotides, followed by their polymerization (Figure 1). However, of the four canonical RNA bases (adenine, cytosine, guanine, uracil), only adenine has been found to react with ribose in a model prebiotic reaction to produce nucleosides in appreciable yields (i.e., about 2%). The other three canonical nucleobases do not produce nucleosides when dried and heated with ribose. This apparent roadblock in RNA synthesis motivated the Orgel laboratory and, more recently, Sutherland and co-workers, to investigate the possibility that the nucleobases were first formed on a pre-existing sugar. -

Monophosphate- Binding Protein Complex with Subsequent Poly(A) RNA Synthesis in Embryonic Chick Cartilage
Specific nuclear binding of adenosine 3',5'-monophosphate- binding protein complex with subsequent poly(A) RNA synthesis in embryonic chick cartilage. W M Burch Jr, H E Lebovitz J Clin Invest. 1980;66(3):532-542. https://doi.org/10.1172/JCI109885. Research Article We used embryonic chick pelvic cartilage as a model to study the mechanism by which cyclic AMP increases RNA synthesis. Isolated nuclei were incubated with [32P]-8-azidoadenosine 3,5'-monophosphate ([32P]N3cAMP) with no resultant specific nuclear binding. However, in the presence of cytosol proteins, nuclear binding of [32P]N3cAMP was demonstrable that was specific, time dependent, and dependent on a heat-labile cytosol factor. The possible biological significance of the nuclear binding of the cyclic AMP-protein complex was identified by incubating isolating nuclei with either cyclic AMP or cytosol cyclic AMP-binding proteins prepared by batch elution DEAE cellulose chromatography (DEAE peak cytosol protein), or both, in the presence of cold nucleotides and [3H]uridine 5'-triphosphate. Poly(A) RNA production occurred only in nuclei incubated with cyclic AMP and the DEAE peak cytosol protein preparation. Actinomycin D inhibited the incorporation of [3H]uridine 5'-monophosphate into poly(A) RNA. The newly synthesized poly(A) RNA had a sedimentation constant of 23S. Characterization of the cytosol cyclic AMP binding proteins using [32P]N3-cAMP with photoaffinity labeling three major cAMP-binding complexes (41,000, 51,000, and 55,000 daltons). The 51,000 and 55,000 dalton cyclic AMP binding proteins were further purified by DNA-cellulose chromatography. In the presence of cyclic AMP they stimulated poly(A) RNA synthesis in isolated nuclei. -

ATP Structure and Function
ATP: Universal Currency of Cellular Energy All living things including plants, animals, birds, insects, humans need energy for the proper functioning of cells, tissues and other organ systems. As we are aware that green plants, obtain their energy from the sunlight, and animals get their energy by feeding on these plants. Energy acts as a source of fuel. We, humans, gain energy from the food we eat, but how are the energy produced and stored in our body. A living cell cannot store significant amounts of free energy. Excess free energy would result in an increase of heat in the cell, which would result in excessive thermal motion that could damage and then destroy the cell. Rather, a cell must be able to handle that energy in a way that enables the cell to store energy safely and release it for use only as needed. Living cells accomplish this by using the compound adenosine triphosphate (ATP). ATP is often called the “energy currency” of the cell, and, like currency, this versatile compound can be used to fill any energy need of the cell. How? It functions similarly to a rechargeable battery. When ATP is broken down, usually by the removal of its terminal phosphate group, energy is released. The energy is used to do work by the cell, usually by the released phosphate binding to another molecule, activating it. For example, in the mechanical work of muscle contraction, ATP supplies the energy to move the contractile muscle proteins. Recall the active transport work of the sodium-potassium pump in cell membranes. -

Inosine in Biology and Disease
G C A T T A C G G C A T genes Review Inosine in Biology and Disease Sundaramoorthy Srinivasan 1, Adrian Gabriel Torres 1 and Lluís Ribas de Pouplana 1,2,* 1 Institute for Research in Biomedicine, Barcelona Institute of Science and Technology, 08028 Barcelona, Catalonia, Spain; [email protected] (S.S.); [email protected] (A.G.T.) 2 Catalan Institution for Research and Advanced Studies, 08010 Barcelona, Catalonia, Spain * Correspondence: [email protected]; Tel.: +34-934034868; Fax: +34-934034870 Abstract: The nucleoside inosine plays an important role in purine biosynthesis, gene translation, and modulation of the fate of RNAs. The editing of adenosine to inosine is a widespread post- transcriptional modification in transfer RNAs (tRNAs) and messenger RNAs (mRNAs). At the wobble position of tRNA anticodons, inosine profoundly modifies codon recognition, while in mRNA, inosines can modify the sequence of the translated polypeptide or modulate the stability, localization, and splicing of transcripts. Inosine is also found in non-coding and exogenous RNAs, where it plays key structural and functional roles. In addition, molecular inosine is an important secondary metabolite in purine metabolism that also acts as a molecular messenger in cell signaling pathways. Here, we review the functional roles of inosine in biology and their connections to human health. Keywords: inosine; deamination; adenosine deaminase acting on RNAs; RNA modification; translation Citation: Srinivasan, S.; Torres, A.G.; Ribas de Pouplana, L. Inosine in 1. Introduction Biology and Disease. Genes 2021, 12, 600. https://doi.org/10.3390/ Inosine was one of the first nucleobase modifications discovered in nucleic acids, genes12040600 having been identified in 1965 as a component of the first sequenced transfer RNA (tRNA), tRNAAla [1]. -

Induced Structural Changes in a Multifunctional Sialyltransferase
Biochemistry 2006, 45, 2139-2148 2139 Cytidine 5′-Monophosphate (CMP)-Induced Structural Changes in a Multifunctional Sialyltransferase from Pasteurella multocida†,‡ Lisheng Ni,§ Mingchi Sun,§ Hai Yu,§ Harshal Chokhawala,§ Xi Chen,*,§ and Andrew J. Fisher*,§,| Department of Chemistry and the Section of Molecular and Cellular Biology, UniVersity of California, One Shields AVenue, DaVis, California 95616 ReceiVed NoVember 23, 2005; ReVised Manuscript ReceiVed December 19, 2005 ABSTRACT: Sialyltransferases catalyze reactions that transfer a sialic acid from CMP-sialic acid to an acceptor (a structure terminated with galactose, N-acetylgalactosamine, or sialic acid). They are key enzymes that catalyze the synthesis of sialic acid-containing oligosaccharides, polysaccharides, and glycoconjugates that play pivotal roles in many critical physiological and pathological processes. The structures of a truncated multifunctional Pasteurella multocida sialyltransferase (∆24PmST1), in the absence and presence of CMP, have been determined by X-ray crystallography at 1.65 and 2.0 Å resolutions, respectively. The ∆24PmST1 exists as a monomer in solution and in crystals. Different from the reported crystal structure of a bifunctional sialyltransferase CstII that has only one Rossmann domain, the overall structure of the ∆24PmST1 consists of two separate Rossmann nucleotide-binding domains. The ∆24PmST1 structure, thus, represents the first sialyltransferase structure that belongs to the glycosyltransferase-B (GT-B) structural group. Unlike all other known GT-B structures, however, there is no C-terminal extension that interacts with the N-terminal domain in the ∆24PmST1 structure. The CMP binding site is located in the deep cleft between the two Rossmann domains. Nevertheless, the CMP only forms interactions with residues in the C-terminal domain. -

Disclosing the Essentiality of Ribose-5-Phosphate Isomerase B In
www.nature.com/scientificreports OPEN Disclosing the essentiality of ribose-5-phosphate isomerase B in Trypanosomatids Received: 04 January 2016 Joana Faria1,2, Inês Loureiro1,2, Nuno Santarém1,2, Pedro Cecílio1,2, Sandra Macedo-Ribeiro2,3, Accepted: 10 May 2016 Joana Tavares1,2,* & Anabela Cordeiro-da-Silva1,2,4,* Published: 27 May 2016 Ribose-5-phosphate isomerase (RPI) belongs to the non-oxidative branch of the pentose phosphate pathway, catalysing the inter-conversion of D-ribose-5-phosphate and D-ribulose-5-phosphate. Trypanosomatids encode a type B RPI, whereas humans have a structurally unrelated type A, making RPIB worthy of exploration as a potential drug target. Null mutant generation in Leishmania infantum was only possible when an episomal copy of RPIB gene was provided, and the latter was retained both in vitro and in vivo in the absence of drug pressure. This suggests the gene is essential for parasite survival. Importantly, the inability to remove the second allele of RPIB gene in sKO mutants complemented with an episomal copy of RPIB carrying a mutation that abolishes isomerase activity suggests the essentiality is due to its metabolic function. In vitro, sKO promastigotes exhibited no defect in growth, metacyclogenesis or macrophage infection, however, an impairment in intracellular amastigotes’ replication was observed. Additionally, mice infected with sKO mutants rescued by RPIB complementation had a reduced parasite burden in the liver. Likewise, Trypanosoma brucei is resistant to complete RPIB gene removal and mice infected with sKO mutants showed prolonged survival upon infection. Taken together our results genetically validate RPIB as a potential drug target in trypanosomatids. -

Metabolomics Identifies Pyrimidine Starvation As the Mechanism of 5-Aminoimidazole-4-Carboxamide-1- Β-Riboside-Induced Apoptosis in Multiple Myeloma Cells
Published OnlineFirst April 12, 2013; DOI: 10.1158/1535-7163.MCT-12-1042 Molecular Cancer Cancer Therapeutics Insights Therapeutics Metabolomics Identifies Pyrimidine Starvation as the Mechanism of 5-Aminoimidazole-4-Carboxamide-1- b-Riboside-Induced Apoptosis in Multiple Myeloma Cells Carolyne Bardeleben1, Sanjai Sharma1, Joseph R. Reeve3, Sara Bassilian3, Patrick Frost1, Bao Hoang1, Yijiang Shi1, and Alan Lichtenstein1,2 Abstract To investigate the mechanism by which 5-aminoimidazole-4-carboxamide-1-b-riboside (AICAr) induces apoptosis in multiple myeloma cells, we conducted an unbiased metabolomics screen. AICAr had selective effects on nucleotide metabolism, resulting in an increase in purine metabolites and a decrease in pyrimidine metabolites. The most striking abnormality was a 26-fold increase in orotate associated with a decrease in uridine monophosphate (UMP) levels, indicating an inhibition of UMP synthetase (UMPS), the last enzyme in the de novo pyrimidine biosynthetic pathway, which produces UMP from orotate and 5-phosphoribosyl- a-pyrophosphate (PRPP). As all pyrimidine nucleotides can be synthesized from UMP, this suggested that the decrease in UMP would lead to pyrimidine starvation as a possible cause of AICAr-induced apoptosis. Exogenous pyrimidines uridine, cytidine, and thymidine, but not purines adenosine or guanosine, rescued multiple myeloma cells from AICAr-induced apoptosis, supporting this notion. In contrast, exogenous uridine had no protective effect on apoptosis resulting from bortezomib, melphalan, or metformin. Rescue resulting from thymidine add-back indicated apoptosis was induced by limiting DNA synthesis rather than RNA synthesis. DNA replicative stress was identified by associated H2A.X phosphorylation in AICAr-treated cells, which was also prevented by uridine add-back. -

Chem 109 C Bioorganic Compounds
Chem 109 C Bioorganic Compounds Fall 2019 HFH1104 Armen Zakarian Office: Chemistry Bldn 2217 http://labs.chem.ucsb.edu/~zakariangroup/courses.html CLAS Instructor: Dhillon Bhavan [email protected] Midterm 3 stats Average 49.1 St Dev 14.1 Max 87.5 Min 15 test are available outside room 2135 (Chemistry, 2nd floor) in a box, sorted in alphabetical order, by color Final Course Grading Each test will be curved individually to 75% average Lowest midterm will be dropped Scores from 2 best M and the Final will be added Grades will be assigned according to the syllabus 22. Draw all reactions required to convert hexanoic acid to 3 molecules of acetyl CoA through 11 pt the β-oxidation cycles. Name all necessary coenzymes and enzymes. How many molecules of ATP and CO2 will be produced from hexanoic acid after its entire metabolism through all 4 stages ? O O hexanoic acid (hexanoate) Overview OVERVIEW o structures of DNA vs. RNA - ribose o structures of bases: Adenine, Uracil/Thymine, Guanine, Cytosine. “Enol forms” o hydrogen bonding between A-T(U) and G-C. H-donors/acceptors o Base complementarity o RNA strand cleavage assisted by the 2’-OH group in the ribose unit (cyclic PDE) o Deamination: RNA genetic instability o DNA replication o RNA synthesis: transcription. Template strand (read 3’ to 5’). Sense strand and the RNA primary structure (T −> U). o Protein synthesis: translation. mRNA determines the amino acid sequence. tRNAs are amino acid carriers. rRNA - part of ribosomes o no section 26.12, 26.13 DNA, RNA, etc. -

Nucleosides & Nucleotides
Nucleosides & Nucleotides Biochemistry Fundamentals > Genetic Information > Genetic Information NUCLEOSIDE AND NUCLEOTIDES SUMMARY NUCLEOSIDES&NBSP; • Comprise a sugar and a base NUCLEOTIDES&NBSP; • Phosphorylated nucleosides (at least one phosphorus group) • Link in chains to form polymers called nucleic acids (i.e. DNA and RNA) N-BETA-GLYCOSIDIC BOND&NBSP; • Links nitrogenous base to sugar in nucleotides and nucleosides • Purines: C1 of sugar bonds with N9 of base • Pyrimidines: C1 of sugar bonds with N1 of base PHOSPHOESTER BOND • Links C3 or C5 hydroxyl group of sugar to phosphate NITROGENOUS BASES&NBSP; • Adenine • Guanine • Cytosine • Thymine (DNA) 1 / 8 • Uracil (RNA) NUCLEOSIDES • =sugar + base • Adenosine • Guanosine • Cytidine • Thymidine • Uridine NUCLEOTIDE MONOPHOSPHATES – ADD SUFFIX 'SYLATE' • = nucleoside + 1 phosphate group • Adenylate • Guanylate • Cytidylate • Thymidylate • Uridylate Add prefix 'deoxy' when the ribose is a deoxyribose: lacks a hydroxyl group at C2. • Thymine only exists in DNA (deoxy prefix unnecessary for this reason) • Uracil only exists in RNA NUCLEIC ACIDS (DNA AND RNA)&NBSP; • Phosphodiester bonds: a phosphate group attached to C5 of one sugar bonds with - OH group on C3 of next sugar • Nucleotide monomers of nucleic acids exist as triphosphates • Nucleotide polymers (i.e. nucleic acids) are monophosphates • 5' end is free phosphate group attached to C5 • 3' end is free -OH group attached to C3 2 / 8 FULL-LENGTH TEXT • Here we will learn about learn about nucleoside and nucleotide structure, and how they create the backbones of nucleic acids (DNA and RNA). • Start a table, so we can address key features of nucleosides and nucleotides. • Denote that nucleosides comprise a sugar and a base.