Molecular Analysis: a New Look at Umbrella Magnolias Richard B
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Foundation Arboretum Wespelaar Year Report 2019
Foundation Arboretum Wespelaar Year Report 2019 2019 started with an early spring: by mid-March the early magnolias were in full flower. Temperatures kept rising and the heat record was broken in July, making the summer of 2019 one to be recorded in history books. Luckily, small rain showers and increased irrigation capacities ensured that we lost very few plants. In March 2019, the construction of the Artois Pavilion started in the Artois Meadow, at the end of two important vistas. The design is inspired by the old pavilion along the canal in the Park of Wespelaar. In the new Arboretum de Marche, a total of 189 trees have been planted in the past two years. In collaboration with Natagora, five ponds were created and a 5ha plot was sown with wild seed to create a meadow of native grasses and flowers. The building permit for the visitors centre and technical facilities was obtained. 2019 was a very active year for both Arboretum Wespelaar and Arboretum de Marche, as is described in more detail in this Year Report. Arboretum Wespelaar – Year Report 2019 1 THE COLLECTIONS ‘Patty’ can be planted in the Arboretum in due time. The living collection of woody plants in the Arboretum currently (as of 30 January 2019) Once again, our winter was not worth contains 5,116 specimens representing 2,340 mentioning and by mid-March we could have different taxa (versus 16,376 specimens and opened the Arboretum for our visitors because 4,955 taxa on the whole of the estate). These the early magnolias were already in full flower! numbers include the 629 new accessions on On March 11th however, we had a serious the estate during 2019 of which 108 (or 17%) storm with accompanying damage and much are of documented wild origin. -

Likely to Have Habitat Within Iras That ALLOW Road
Item 3a - Sensitive Species National Master List By Region and Species Group Not likely to have habitat within IRAs Not likely to have Federal Likely to have habitat that DO NOT ALLOW habitat within IRAs Candidate within IRAs that DO Likely to have habitat road (re)construction that ALLOW road Forest Service Species Under NOT ALLOW road within IRAs that ALLOW but could be (re)construction but Species Scientific Name Common Name Species Group Region ESA (re)construction? road (re)construction? affected? could be affected? Bufo boreas boreas Boreal Western Toad Amphibian 1 No Yes Yes No No Plethodon vandykei idahoensis Coeur D'Alene Salamander Amphibian 1 No Yes Yes No No Rana pipiens Northern Leopard Frog Amphibian 1 No Yes Yes No No Accipiter gentilis Northern Goshawk Bird 1 No Yes Yes No No Ammodramus bairdii Baird's Sparrow Bird 1 No No Yes No No Anthus spragueii Sprague's Pipit Bird 1 No No Yes No No Centrocercus urophasianus Sage Grouse Bird 1 No Yes Yes No No Cygnus buccinator Trumpeter Swan Bird 1 No Yes Yes No No Falco peregrinus anatum American Peregrine Falcon Bird 1 No Yes Yes No No Gavia immer Common Loon Bird 1 No Yes Yes No No Histrionicus histrionicus Harlequin Duck Bird 1 No Yes Yes No No Lanius ludovicianus Loggerhead Shrike Bird 1 No Yes Yes No No Oreortyx pictus Mountain Quail Bird 1 No Yes Yes No No Otus flammeolus Flammulated Owl Bird 1 No Yes Yes No No Picoides albolarvatus White-Headed Woodpecker Bird 1 No Yes Yes No No Picoides arcticus Black-Backed Woodpecker Bird 1 No Yes Yes No No Speotyto cunicularia Burrowing -

Indiana's Native Magnolias
FNR-238 Purdue University Forestry and Natural Resources Know your Trees Series Indiana’s Native Magnolias Sally S. Weeks, Dendrologist Department of Forestry and Natural Resources Purdue University, West Lafayette, IN 47907 This publication is available in color at http://www.ces.purdue.edu/extmedia/fnr.htm Introduction When most Midwesterners think of a magnolia, images of the grand, evergreen southern magnolia (Magnolia grandiflora) (Figure 1) usually come to mind. Even those familiar with magnolias tend to think of them as occurring only in the South, where a more moderate climate prevails. Seven species do indeed thrive, especially in the southern Appalachian Mountains. But how many Hoosiers know that there are two native species Figure 2. Cucumber magnolia when planted will grow well throughout Indiana. In Charles Deam’s Trees of Indiana, the author reports “it doubtless occurred in all or nearly all of the counties in southern Indiana south of a line drawn from Franklin to Knox counties.” It was mainly found as a scattered, woodland tree and considered very local. Today, it is known to occur in only three small native populations and is listed as State Endangered Figure 1. Southern magnolia by the Division of Nature Preserves within Indiana’s Department of Natural Resources. found in Indiana? Very few, I suspect. No native As the common name suggests, the immature magnolias occur further west than eastern Texas, fruits are green and resemble a cucumber so we “easterners” are uniquely blessed with the (Figure 3). Pioneers added the seeds to whisky presence of these beautiful flowering trees. to make bitters, a supposed remedy for many Indiana’s most “abundant” species, cucumber ailments. -

Download PCN Magnolia Multisite
Institution name plant NAMES for inventory::print name Accession # Provenanc Quantity Plant source The Scott Arboretum atMagnolia Swarthmore acuminata College 2005-355UN*A G 1 Unknown The Scott Arboretum atMagnolia Swarthmore acuminata College 2001-188UN*A U 1 Unknown The Scott Arboretum atMagnolia Swarthmore acuminata College 96-129*A G 1 Princeton Nurseries The Scott Arboretum atMagnolia Swarthmore acuminata College var. subcordata 99-203*B G 1 Longwood Gardens The Scott Arboretum atMagnolia Swarthmore acuminata College var. subcordata 93-206*A G 1 Woodlanders Nursery The Scott Arboretum atMagnolia Swarthmore acuminata College var. subcordata 'Brenda'2004-239*A G 1 Pat McCracken The Scott Arboretum atMagnolia Swarthmore 'Anilou' College 2008-202*A G 1 Pleasant Run Nursery The Scott Arboretum atMagnolia Swarthmore 'Anilou' College 2008-202*B G 1 Pleasant Run Nursery The Scott Arboretum atMagnolia Swarthmore 'Ann' College 68-165*A G 1 U. S. National Arboretum, Washington, DC The Scott Arboretum atMagnolia Swarthmore 'Banana College Split' 2004-237*A G 1 Pat McCracken The Scott Arboretum atMagnolia Swarthmore 'Betty' College 68-166*A G 1 U. S. National Arboretum, Washington, DC The Scott Arboretum atMagnolia Swarthmore 'Big Dude' College 2008-203*A G 1 Pleasant Run Nursery The Scott Arboretum atMagnolia Swarthmore ×brooklynensis College 'Black Beauty' 2008-204*A G 1 Pleasant Run Nursery The Scott Arboretum atMagnolia Swarthmore ×soulangeana College 'Jurmag1' 2010-069*A G 1 Pleasant Run Nursery The Scott Arboretum atMagnolia Swarthmore -

THE Magnoliaceae Liriodendron L. Magnolia L
THE Magnoliaceae Liriodendron L. Magnolia L. VEGETATIVE KEY TO SPECIES IN CULTIVATION Jan De Langhe (1 October 2014 - 28 May 2015) Vegetative identification key. Introduction: This key is based on vegetative characteristics, and therefore also of use when flowers and fruits are absent. - Use a 10× hand lens to evaluate stipular scars, buds and pubescence in general. - Look at the entire plant. Young specimens, shade, and strong shoots give an atypical view. - Beware of hybridisation, especially with plants raised from seed other than wild origin. Taxa treated in this key: see page 10. Questionable/frequently misapplied names: see page 10. Names referred to synonymy: see page 11. References: - JDL herbarium - living specimens, in various arboreta, botanic gardens and collections - literature: De Meyere, D. - (2001) - Enkele notities omtrent Liriodendron tulipifera, L. chinense en hun hybriden in BDB, p.23-40. Hunt, D. - (1998) - Magnolias and their allies, 304p. Bean, W.J. - (1981) - Magnolia in Trees and Shrubs hardy in the British Isles VOL.2, p.641-675. - or online edition Clarke, D.L. - (1988) - Magnolia in Trees and Shrubs hardy in the British Isles supplement, p.318-332. Grimshaw, J. & Bayton, R. - (2009) - Magnolia in New Trees, p.473-506. RHS - (2014) - Magnolia in The Hillier Manual of Trees & Shrubs, p.206-215. Liu, Y.-H., Zeng, Q.-W., Zhou, R.-Z. & Xing, F.-W. - (2004) - Magnolias of China, 391p. Krüssmann, G. - (1977) - Magnolia in Handbuch der Laubgehölze, VOL.3, p.275-288. Meyer, F.G. - (1977) - Magnoliaceae in Flora of North America, VOL.3: online edition Rehder, A. - (1940) - Magnoliaceae in Manual of cultivated trees and shrubs hardy in North America, p.246-253. -

Magnolia Macrophylla: Bigleaf Magnolia1 Edward F
ENH-540 Magnolia macrophylla: Bigleaf Magnolia1 Edward F. Gilman and Dennis G. Watson2 Introduction green above with a fuzzy, silver/grey underside, creating a beautiful, two-toned effect with each passing breeze. From This North American native tree is deciduous in most areas May to July the showy, fragrant blossoms appear, each 8 to but semi-evergreen in the Deep South. Bigleaf Magnolia 12-inch-wide, ivory-colored bloom having a slight rose tint grows slowly to 30 to 40 feet and spreads 20 to 25 feet at its base. These blooms are followed by the production forming a rounded, broad canopy. The leaves of Bigleaf of 2.5 to 3-inch-long, hairy, red, egg-shaped fruits. Bigleaf Magnolia are truly large, 12 to 32 inches long and 7 to Magnolia trees must be 12 to 15-years-of-age before they 12 inches wide, when found in the wild and somewhat begin to bloom. smaller when grown in landscapes. These leaves are bright General Information Scientific name: Magnolia macrophylla Pronunciation: mag-NO-lee-uh mack-roe-FILL-uh Common name(s): Bigleaf Magnolia Family: Magnoliaceae USDA hardiness zones: 5B through 8B (Fig. 2) Origin: native to North America Figure 1. Young Magnolia macrophylla: Bigleaf Magnolia Credits: Ed Gilman Figure 2. Range 1. This document is ENH-540, one of a series of the Environmental Horticulture, UF/IFAS Extension. Original publication date November 1993. Revised December 2006. Reviewed February 2014. Visit the EDIS website at http://edis.ifas.ufl.edu. 2. Edward F. Gilman, professor, Environmental Horticulture Department; Dennis G. -

Meristems West Tisbury, Massachusetts Vol
The Polly Hill Arboretum Meristems West Tisbury, Massachusetts Vol. 16, No. 2 Fall 2014 Ilex opaca ‘Villanova’: a yellow-fruited American holly selected and named by Polly Hill. Education Center Receives Matching Grant PHA continues to grow at a measured Situated at the heart of the Arboretum qualities of plants inspires our day-to-day pace. In 2006 the Arboretum outlined sev - campus between the Homestead (our work. Coupled with this is our desire eral capital projects to advance our mission administrative offices) and the Cowbarn, to share our enthusiasm and knowledge of of education, horticultural experimentation, the Education Center and Botany Lab plants with a larger group of children and plant conservation. We have been will provide a climate-controlled indoor and adults. The proposed building helps successful with a new greenhouse (2006), environment to extend our education us accomplish all these goals. the Cowbarn renovation (2007), the programming year-round and the space and Now the amazing news! PHA has Littlefield Maintenance Building (2009), equipment to advance our plant research. received a $500,000 gift from the Cedar and a refurbished Far Barn (2011). Its central location is visible from State Tree Foundation (the family foundation Today one significant project remains: Road, making its design and positioning of our founder, Dr. David Smith) to raise the proposed Education Center and Botany critical to maintaining the spirit of our matching funds for this new building. Lab. We have completed plans for this historical landscape. This generous grant supports our most building slated to replace the dilapidated PHA staff and board of directors are important education and scientific building outbuilding known as the Gym. -
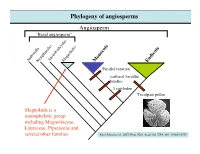
Phylogeny of Angiosperms Angiosperm “Basal Angiosperm”
Phylogeny of angiosperms Angiosperm “Basal angiosperm” AmborellaNymphaealesAustrobaileyalesMagnoliidss Monocots Eudicots Parallel venation scattered vascular bundles 1 cotyledon Tricolpate pollen Magnoliids is a monophyletic group including Magnoliaceae, Lauraceae, Piperaceae and several other families After Jansen et al., 2007, Proc. Natl. Acad. Sci. USA 104: 19369-19374 Magnoliaceae (Magnolia family) Textbook DVD KRR Magnolia X soulangeana; Magnoliaceae (Magnolia family) Textbook DVD WSJ Textbook DVD KRR Magnolia grandiflora; Magnolia macrophylla; Note leaf simple, entire, pinnate venation, numerous tepals, numerous stamens and carples. Textbook DVD KRR Magnolia sieboldii; Magnoliaceae (Magnolia family) Textbook DVD KMN Textbook DVD SMK-KRR Magnolia figo; Magnolia grandiflora; Note the elongated receptacle, Note the aggregate of follicles, and laminar stamens and red fleshy seed coat Magnoliaceae (Magnolia family) Photo: Yaowu Yuan Photo: Yaowu Yuan Liriodendron tulipifera; Note the elongated receptacle, and laminar stamens Magnoliaceae (Magnolia family) Note the lobed, T-shirt-like leaf, and pinnate venation Note the aggregate of samara Magnoliaceae (Magnolia family) Magnoliaceae - 2 genera/220 species. Trees or shrubs; Ethereal oils (aromatic terpenoids) - (remember the smell of bay leaves?); Leaves alternate, simple (Magnolia) or lobed (Liriodendron), entire; Flowers large and showy, actinomorphic, bisexual Tepals 6-numerous, stamens and carpels numerous, Spirally arranged on an elongated receptacle, Laminar stamens poorly differentiated into anther and filament. Fruit usually an aggregate of follicle (Magnolia) or samara (Liriodendron); follicle: 1-carpellate fruit that dehisces on the side samara: 1-carpellate winged, indehiscent fruit Phylogeny of Eudicots (or Tricolpates) Eudicots (or Tricolpates) “Basal eudicots” Asterids Buxales Rosids Caryophyllales RanunculalesProteales Ranunculales is a monophyletic group including Ranunculaceae, Berberidaceae, Papaveraceae, and 4 other families. After Jansen et al., 2007, Proc. -

Reconstructing the Basal Angiosperm Phylogeny: Evaluating Information Content of Mitochondrial Genes
55 (4) • November 2006: 837–856 Qiu & al. • Basal angiosperm phylogeny Reconstructing the basal angiosperm phylogeny: evaluating information content of mitochondrial genes Yin-Long Qiu1, Libo Li, Tory A. Hendry, Ruiqi Li, David W. Taylor, Michael J. Issa, Alexander J. Ronen, Mona L. Vekaria & Adam M. White 1Department of Ecology & Evolutionary Biology, The University Herbarium, University of Michigan, Ann Arbor, Michigan 48109-1048, U.S.A. [email protected] (author for correspondence). Three mitochondrial (atp1, matR, nad5), four chloroplast (atpB, matK, rbcL, rpoC2), and one nuclear (18S) genes from 162 seed plants, representing all major lineages of gymnosperms and angiosperms, were analyzed together in a supermatrix or in various partitions using likelihood and parsimony methods. The results show that Amborella + Nymphaeales together constitute the first diverging lineage of angiosperms, and that the topology of Amborella alone being sister to all other angiosperms likely represents a local long branch attrac- tion artifact. The monophyly of magnoliids, as well as sister relationships between Magnoliales and Laurales, and between Canellales and Piperales, are all strongly supported. The sister relationship to eudicots of Ceratophyllum is not strongly supported by this study; instead a placement of the genus with Chloranthaceae receives moderate support in the mitochondrial gene analyses. Relationships among magnoliids, monocots, and eudicots remain unresolved. Direct comparisons of analytic results from several data partitions with or without RNA editing sites show that in multigene analyses, RNA editing has no effect on well supported rela- tionships, but minor effect on weakly supported ones. Finally, comparisons of results from separate analyses of mitochondrial and chloroplast genes demonstrate that mitochondrial genes, with overall slower rates of sub- stitution than chloroplast genes, are informative phylogenetic markers, and are particularly suitable for resolv- ing deep relationships. -

Ouachita Mountains Ecoregional Assessment December 2003
Ouachita Mountains Ecoregional Assessment December 2003 Ouachita Ecoregional Assessment Team Arkansas Field Office 601 North University Ave. Little Rock, AR 72205 Oklahoma Field Office 2727 East 21st Street Tulsa, OK 74114 Ouachita Mountains Ecoregional Assessment ii 12/2003 Table of Contents Ouachita Mountains Ecoregional Assessment............................................................................................................................i Table of Contents ........................................................................................................................................................................iii EXECUTIVE SUMMARY..............................................................................................................1 INTRODUCTION..........................................................................................................................3 BACKGROUND ...........................................................................................................................4 Ecoregional Boundary Delineation.............................................................................................................................................4 Geology..........................................................................................................................................................................................5 Soils................................................................................................................................................................................................6 -

System for Satellite-Based Classification of Plant
ecologies Article Genus-Physiognomy-Ecosystem (GPE) System for Satellite-Based Classification of Plant Communities Ram C. Sharma Department of Informatics, Tokyo University of Information Sciences, 4-1 Onaridai, Wakaba, Chiba 265-8501, Japan; [email protected]; Tel.: +81-43-236-4603 Abstract: Vegetation mapping and monitoring is important as the composition and distribution of vegetation has been greatly influenced by land use change and the interaction of land use change and climate change. The purpose of vegetation mapping is to discover the extent and distribution of plant communities within a geographical area of interest. The paper introduces the Genus-Physiognomy- Ecosystem (GPE) system for the organization of plant communities from the perspective of satellite remote sensing. It was conceived for broadscale operational vegetation mapping by organizing plant communities according to shared genus and physiognomy/ecosystem inferences, and it offers an intermediate level between the physiognomy/ecosystem and dominant species for the organi- zation of plant communities. A machine learning and cross-validation approach was employed by utilizing multi-temporal Landsat 8 satellite images on a regional scale for the classification of plant communities at three hierarchical levels: (i) physiognomy, (ii) GPE, and (iii) dominant species. The classification at the dominant species level showed many misclassifications and undermined its application for broadscale operational mapping, whereas the GPE system was able to lessen the complexities associated with the dominant species level classification while still being capable of distinguishing a wider variety of plant communities. The GPE system therefore provides an easy-to-understand approach for the operational mapping of plant communities, particularly on a Citation: Sharma, R.C. -

The Wonderful Ashe Magnolia
The wonderful Ashe magnolia Charles E. Selter I have an Ashe magnoha growing on a street corner at my residence in Taflahassee, Florida. When it blooms cars stop suddenly and pull over to the curb. People get out of the car, walk up to the tree, and stare. I can accurately say Ashe magnolia stops trafflc( It looks and smells great. Part of the reaction to my tree is because you don't see an Ashe magnofla on every street corner. In fact. you don't see them often anywhere, which is a great pity because they have great horticultural value. 'Ihe foliage of Ashe magnoha (Magnolia maerophylia var. asheij is attractive. The leaves mature at 12 to 16 inches in length. Leaves are 6 to 8 inches wide above midpoint, tapered to their base with a rounded ear on each side of the midrib (auriculate). The apex of the leaf is rounded with a pointed tip. 'Ihe leaf is thin and flexible tn a breeze, deep green on top and whitish on their undersides. 'Ihe petiole is 4 to 5 inches long. stout, and extends as a midrib to the tip of the leaf. 'Ihe deep summer green tume to yellow in the fall and, after falBng, the upper surface tume a chocolate brown whfle the lower surface tume sflvery-white. Fallen leaves are beautiful and unusual in autumn leaf floral arrangements. 'Ihe leaves sre home in a whorl at the end of branches snd twigs and thus form an open crown of ezcttc texts. The flowers of Ashe magnofla are large, spectacular, iragrant and numerous.