ISOLATION of Bacillus Thuringiensis and ITS EFFECT AGAINST Galleria Mellonella
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Attract-And-Kill Methods for Control of Indianmeal Moth
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by SHAREOK repository ATTRACT-AND-KILL METHODS FOR CONTROL OF INDIANMEAL MOTH, Plodia interpunctella (Hübner) (LEPIDOPTERA: PYRALIDAE), AND COMPARISONS WITH OTHER PHEROMONE-BASED CONTROL METHODS By MANUEL CAMPOS-FIGUEROA Bachelor of Science in Entomology and Plant Pathology Universidad Autónoma Chapingo Chapingo, State of México, México 1996 Master of Science in Entomology Colegio de Postgraduados Montecillos, State of México, México 1999 Submitted to the Faculty of the Graduate College of the Oklahoma State University in partial fulfillment of the requirements for the Degree of DOCTOR OF PHILOSOPHY May, 2009 ATTRACT-AND-KILL METHODS FOR CONTROL OF INDIANMEAL MOTH, Plodia interpunctella (Hübner) (LEPIDOPTERA: PYRALIDAE), AND COMPARISONS WITH OTHER PHEROMONE-BASED CONTROL METHODS Dissertation Approved: Dr. Thomas W. Phillips Dissertation Adviser Dr. Mark E. Payton Dr. Jack W. Dillwith Dr. Brad Kard Dr. A. Gordon Emslie Dean of the Graduate College ii PREFACE I want to extend my gratitude to my major advisor and mentor Dr. Thomas W. Phillips for his time and support given to me during my Ph.D. studies. Also, I appreciate Dr. Phillips for considering me part of your research team. You are an example to follow, a great scientist, a great person and always looking for a solution. I am grateful with Dr. Jack W. Dillwith for being an excellent professor and committee member. Dr. Dillwith has always been very helpful and gave me good suggestions that improved this research. Thank you for being such a great support during all this time. -

Acoustic Communication in the Nocturnal Lepidoptera
Chapter 6 Acoustic Communication in the Nocturnal Lepidoptera Michael D. Greenfield Abstract Pair formation in moths typically involves pheromones, but some pyra- loid and noctuoid species use sound in mating communication. The signals are generally ultrasound, broadcast by males, and function in courtship. Long-range advertisement songs also occur which exhibit high convergence with commu- nication in other acoustic species such as orthopterans and anurans. Tympanal hearing with sensitivity to ultrasound in the context of bat avoidance behavior is widespread in the Lepidoptera, and phylogenetic inference indicates that such perception preceded the evolution of song. This sequence suggests that male song originated via the sensory bias mechanism, but the trajectory by which ances- tral defensive behavior in females—negative responses to bat echolocation sig- nals—may have evolved toward positive responses to male song remains unclear. Analyses of various species offer some insight to this improbable transition, and to the general process by which signals may evolve via the sensory bias mechanism. 6.1 Introduction The acoustic world of Lepidoptera remained for humans largely unknown, and this for good reason: It takes place mostly in the middle- to high-ultrasound fre- quency range, well beyond our sensitivity range. Thus, the discovery and detailed study of acoustically communicating moths came about only with the use of electronic instruments sensitive to these sound frequencies. Such equipment was invented following the 1930s, and instruments that could be readily applied in the field were only available since the 1980s. But the application of such equipment M. D. Greenfield (*) Institut de recherche sur la biologie de l’insecte (IRBI), CNRS UMR 7261, Parc de Grandmont, Université François Rabelais de Tours, 37200 Tours, France e-mail: [email protected] B. -

Managing Maladies: Parasites, Pests & Pesticides
MANAGING MALADIES: PARASITES, PESTS & PESTICIDES W. Brian Kreowski Agricultural Inspector Today’s Objectives: • Briefly explain the Virginia Apiary Inspection program • Go over the main insects and mites that might harm your bees • Discuss animals that can potentially harm your hive(s) and their contents • Make you aware of the hazards of pesticides to your investment in bees Apiary Inspection Program • Inspector’s duties: • Examine bees for disease, etc. • Prohibit movement or sale of diseased hives • Destroy hives when necessary Apiary Inspection Program • Beekeepers role: • Provide movable frames • Tightly close dead hives • Notify State Apiarist of diseased bees • Notify State Apiarist of possible Africanized Honeybee Apiary Inspection Program: Regulations – No honey in candy for queen cages – Queen rearing and queen mating apiary inspection – Inspection certificate for packages – Inspection prior to bringing hives into Commonwealth – Certificate for sale of comb, hives, used equipment with comb or appliances Parasites: Varroa Mite • Varroa destructor Anderson & Trueman • External parasite of adults… Parasites: Varroa Mite • …and immatures Parasites: Varroa Mite Parasites: Varroa Mite - Biology • Female mites crawl into brood cells (especially drone cells) before they are capped • Feed on bee food first, then on the prepupa • Lay the first eggs about 60 hours after cell capping, subsequent eggs are laid at 30 hour intervals • Lay about 4 to 6 eggs total Parasites: Varroa Mite - Biology • Immature mites develop on the pupal bee, requiring -

Beekeepers and Bee Wax Moth
Faculty of Natural Resources and Agricultural Sciences Beekeepers and Bee Wax Moth – What’s the Buzz About? A study about the attitude towards beekeeping and wax moth in Kenya Biodlare och bivaxmal – vad är allt surrande om? – En studie om attityden mot biodling och bivaxmal i Kenya Ida Johansson Independent project in Biology • 15 hec Agriculture program - plant and soil sciences Department of Ecology Uppsala 2019 Beekeepers and Bee Wax Moth – What’s the Buzz About? A study about the attitude towards beekeeping and wax moth in Kenya Biodlare och bivaxmal – vad är allt surrande om? – En studie om attityden mot biodling och bivaxmal i Kenya Ida Johansson Supervisor: Helena Bylund, SLU, Department of Ecology Assistant supervisor: Jamleck Muturi, University of Embu, Department of Biological Sciences Examiner: Mattias Jonsson, SLU, Department of Ecology Credits: 15 hec Level: G2E Course title: Independent project in Biology Course code: EX0894 Programme/education: Agriculture Program – Plant and Soil Sciences Course coordinating department: Department of Aquatic Sciences and Assessment Place of publication: Uppsala Year of publication: 2019 Cover picture: Ida Johansson Online publication: https://stud.epsilon.slu.se Keywords: Pollination, wax moth, Kenya, bees, beekeeping Sveriges lantbruksuniversitet Swedish University of Agricultural Sciences Faculty of natural resources and Agricultural Sciences Department of Ecology Abstract The bee wax moth (Galleria mellonella) is a troublesome pest of the honeybees that can cause substantial financial losses that can be devastating for the beekeepers. The moth is a universal pest but thrives in a warmer climate, the insect is nocturnal and feed on pollen, wax and other impurities. The eggs are laid in crevice’s and other small spaces where the bees cannot reach them. -

Moth Hearing and Sound Communication
J Comp Physiol A (2015) 201:111–121 DOI 10.1007/s00359-014-0945-8 REVIEW Moth hearing and sound communication Ryo Nakano · Takuma Takanashi · Annemarie Surlykke Received: 17 July 2014 / Revised: 13 September 2014 / Accepted: 15 September 2014 / Published online: 27 September 2014 © Springer-Verlag Berlin Heidelberg 2014 Abstract Active echolocation enables bats to orient notion of moths predominantly being silent. Sexual sound and hunt the night sky for insects. As a counter-measure communication in moths may apply to many eared moths, against the severe predation pressure many nocturnal perhaps even a majority. The low intensities and high fre- insects have evolved ears sensitive to ultrasonic bat calls. quencies explain that this was overlooked, revealing a bias In moths bat-detection was the principal purpose of hear- towards what humans can sense, when studying (acoustic) ing, as evidenced by comparable hearing physiology with communication in animals. best sensitivity in the bat echolocation range, 20–60 kHz, across moths in spite of diverse ear morphology. Some Keywords Co-evolution · Sensory exploitation · eared moths subsequently developed sound-producing Ultrasound · Echolocating bats · Predator-prey organs to warn/startle/jam attacking bats and/or to com- municate intraspecifically with sound. Not only the sounds for interaction with bats, but also mating signals are within Introduction the frequency range where bats echolocate, indicating that sound communication developed after hearing by “sensory Insect hearing and sound communication is a fascinating exploitation”. Recent findings on moth sound communica- subject, where the combination of many classical studies tion reveal that close-range (~ a few cm) communication and recent progress using new technological and molecular with low-intensity ultrasounds “whispered” by males dur- methods provide an unsurpassed system for studying and ing courtship is not uncommon, contrary to the general understanding the evolution of acoustic communication, intraspecific as well as between predator and prey. -

4 Biology, Behavior, and Ecology of Insects in Processed Commodities
4 Biology, Behavior, and Ecology of Insects in Processed Commodities Rizana M. Mahroof David W. Hagstrum Most insects found in storage facilities consume Red flour beetle, Tribolium commodities, but some feed on mold growing castaneum (Herbst) on stored products. Others may be predators and parasitoids. Insects that attack relatively dry pro- Red flour beetle adults (Figure 1) are reddish brown. cessed commodities (those with about 10% or more Eggs are oblong and white. Adults show little moisture content at 15 to 42oC) can cause signifi- preference for cracks or crevices as oviposition sites. cant weight losses during storage. Insects occur in Eggshells are coated with a sticky substance that aids flour mills, rice mills, feed mills, food processing in attaching the eggs to surfaces and causes small facilities, breakfast and cereal processing facilities, particles to adhere to them (Arbogast 1991). Larvae farm storages, grain bins, grain elevators, bakeries, are yellowish white with three pair of thoracic legs. warehouses, grocery stores, pet-food stores, herbari- ums, museums, and tobacco curing barns. Economic Typically, there are six to seven larval instars, losses attributed to insects include not only weight depending on temperature and nutrition. Larvae loss of the commodity, but also monitoring and pest move away from light, living concealed in the food. management costs and effects of contamination on Full-grown larvae move to the food surface or seek product trade name reputation. shelter for pupation. Pupae are white and exarate, which means that appendages are not fused to the body. External genitalic characters on pupae can be Life Histories used to differentiate males and females (Good 1936). -
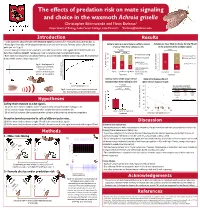
Results Methods Introduction Christopher Edomwande and Flavia Barbosa1 Hypotheses Discussion References Acknowledgements
The effects of predation risk on mate signaling and choice in the waxmoth Achroia grisella Christopher Edomwande and Flavia Barbosa1 Department of Biology, Lake Forest College, Lake Forest IL [email protected] Introduction Results • Male waxmoths attract females with ultrasonic signals consisting of an incessant pulse pair train (fig. 1). • Males signal from leks, which receptive females orient and walk towards. Females prefer calls with faster Solitary males are more likely to exhibit a silence Females are more likely to choose the low PR call pulse‐pair rate1 (fig. 2). response than those calling in a lek. in the presence of the predator signal. • Several mating behaviors make waxmoths vulnerable to predators: male signals attract both females and **p<0.001 *p=0.03 bats, thus creating a tradeoff. Females are more vulnerable when moving towards males. *p=0.014 p=0.06 20 Continuous callingSeries2 12 • Both sexes may respond to bat calls by freezing, and calling males exhibit a silence response. More attractive 2 Chose low PR call males exhibit shorter silence responses2‐4 . 15 8 Silence responseSeries1 9 8 Chose high PR call 13 10 Cochran Q test: 6 13 17 2 Fig. 1 – Oscillogram of a X2 = 15.1667, p<0.001. McNemar test: X = 4.5 5 11 3 typical male call. Males Number of males 5 6 Pairwise comparisons: typically produce 0 McNemar test, Number of females 0 between 70 to 110 pulse Silence Low PR lek High PR lek Bonferroni adjusted No predator Predator pairs s‐1, at 70–130 kHz. Treatment alpha = 0.016. -

Indianmeal Moth, Plodia Interpunctella (Hübner) (Insecta: Lepidoptera: Pyralidae)1 Thomas R
EENY-026 Indianmeal Moth, Plodia interpunctella (Hübner) (Insecta: Lepidoptera: Pyralidae)1 Thomas R. Fasulo and Marle A. Knox2 Introduction Larvae The Indianmeal moth, Plodia interpunctella (Hübner), is a There are five to seven larval instars. Their color is usually very common household pest, feeding principally on stored off-white, but has been observed to be pink, brown, or food products. In fact, it has been called the most impor- almost greenish, depending on the food source. The mature tant pest of stored products commonly found in American larvae are about 1.27 cm in length. They have five pairs of homes or grocery stores. The larvae are general feeders, well-developed prolegs that help them move considerable which may feed on grain products, seeds, dried fruit, dog distances to pupate. food, and spices. The Indianmeal moth received its com- mon name from the United States where it was found to be a pest of meal made of “Indian corn” or maize. Distribution This insect is found in a wide range of climates in stored products and food storage facilities around the world. It is very common in Florida, where it also lives successfully outdoors. Description Eggs Eggs of the Indianmeal moth appear grayish-white and range in length from 0.3 to 0.5 mm. Eggs are oviposited singly or in clusters, and are generally laid directly on the larval food source. Figure 1. Larva of the Indianmeal moth, Plodia interpunctella (Hübner). Credits: Lyle Buss, UF/IFAS 1. This document is EENY-026, one of a series of the Department of Entomology and Nematology, UF/IFAS Extension. -

Stored Product Pests Department of Entomology
PURDUE EXTENSION E-37-W Stored Product Pests Department of Entomology INSECT PESTS OF HOME STORED FOODS Linda J. Mason and Timothy J. Gibb, Extension Entomologists Many kinds of cereal products and other foods stored GRAIN AND FLOUR BEETLES in kitchen cabinets or elsewhere in the home may become Sometimes collectively called "bran bugs," these infested with insects or other organisms commonly reddish-brown beetles are usually less than 1/8 inch referred to as "pantry pests." Practically all dried food long. Their small, wormlike larvae (growing stages) are products commonly found in the home are susceptible, yellowish-white with brown heads. Larvae of the first four including birdseed and dry pet foods. Pantry pests eat or species illustrated are elongate and tubular; those of the contaminate food, thus making it unfit for humans. They latter two species are somewhat C-shaped in appearance may also be annoying, in that they often leave the infested and appear rather hairy. The larvae are usually found in food and crawl or fly about the house. To eliminate infested material, whereas adult beetles often crawl about infestations, it is necessary to identify the pest and then the kitchen or other areas as well as feed in the infested find and destroy or treat infested materials. Listed and material. illustrated below by groups are the most common pests of stored food products in Indiana. Confused Flour Beetle Cigarette Beetle Drugstore Beetle Red Flour Beetle Saw-Toothed Grain Beetle NOTE: Indicates relative size through this whole publication Insect Pests of Home Stored Foods — E-37-W 2 DERMESTID BEETLES SPIDER BEETLES Members of this family are generally scavengers Several species of spider beetles (long legs and a and feed on a great variety of products of both plant general spider-like appearance) may be found infesting all and animal origin including leather, furs, skins, dried types of stored food products. -

Eb 1396 1994 Meal Moths.Pdf (2.335Mb)
Declared out of print June 2013. Some facts and recommendations in this publication are no longer endorsed by WSU Extension. Please look for up-to-date information in the WSU Extension Online Store at https://pubs.wsu.edu. SCI S tension Bulletin 1396 537 W2573 no.1396 1994 MEAL MOTHS Meal moths attack stored grain products or house Indianmeal Moth hold foodstuffs. Once established in food, insect populations can increase and infest vulnerable ma The adult Indianmeal moth, Plodia interpunctella, terial throughout the home, apartment, or storage has a wingspan of about 1/2 to 3/4-inch. The tips or area. Some adult moths do fly into the home outer half of the wings are reddish brown or "cop through open doors or windows, but most are car pery," while the basal portions are grayish-white. The ried inside from outdoor storage or in packaged larvae are usually off-white but can be pink, yellow, goods or groceries. greenish, or brownish. At maturity the larvae are about 1/2-inch long. Everyone's home is vulnerable. However, those who do not store food properly have the greatest The adult female can lay several dozen to several problems. Spilled or exposed foods attract the in hundred eggs. The larval stage may last from a few sects and increase the chance of infestation. Foods weeks to 9 or 10 months, depending on temperature. that are not tightly sealed, especially those main The larvae, which spin a great deal of webbing while tained for long periods of time, are particularly they are feeding and maturing, are usually found in susceptible to infestation. -

<I>Plodia Interpunctella</I>
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln U.S. Department of Agriculture: Agricultural Publications from USDA-ARS / UNL Faculty Research Service, Lincoln, Nebraska 2007 Biology and management of Plodia interpunctella (Lepidoptera: Pyralidae) in stored products S. Mohandass Kansas State University F. H. Arthur USDA-ARS, [email protected] K. Y. Zhu Kansas State University, [email protected] James E. Throne USDA-ARS, [email protected] Follow this and additional works at: https://digitalcommons.unl.edu/usdaarsfacpub Mohandass, S.; Arthur, F. H.; Zhu, K. Y.; and Throne, James E., "Biology and management of Plodia interpunctella (Lepidoptera: Pyralidae) in stored products" (2007). Publications from USDA-ARS / UNL Faculty. 1981. https://digitalcommons.unl.edu/usdaarsfacpub/1981 This Article is brought to you for free and open access by the U.S. Department of Agriculture: Agricultural Research Service, Lincoln, Nebraska at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Publications from USDA-ARS / UNL Faculty by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. ARTICLE IN PRESS Journal of Stored Products Research 43 (2007) 302–311 www.elsevier.com/locate/jspr Review Biology and management of Plodia interpunctella (Lepidoptera: Pyralidae) in stored products$ S. Mohandassa, F.H. Arthurb,Ã, K.Y. Zhuc, J.E. Throneb aDepartment of Architecture, Planning, and Design, Kansas State University, Manhattan, KS, USA bUSDA-ARS, Grain Marketing and Production Research Center, 1515 College Avenue, Manhattan, KS-66506, USA cDepartment of Entomology, Kansas State University, Manhattan, KS, USA Accepted 10 August 2006 Abstract Plodia interpunctella (Hu¨bner), the Indian meal moth, is a world-wide insect pest of stored-products and processed food commodities. -

Parasites of the Honeybee
Parasites of the Honeybee This publication was produced by Dr. Mary F. Coffey Teagasc, Crops Research Centre, Oak Park, Carlow co-financed by The Department of Agriculture, Fisheries and Food November 2007 Table of Contents Introduction……………………………………………………………………….………….…….1 Bee origin and classification………………..……………………………………………………..2 The honeybee colony……….…………………………………..……………………......................4 Irish floral sources of pollen and nectar……………………………………...…………….…….11 Pests and diseases of the honeybee………………………………………………………….……12 Pests……………………………………….………………………………………...…………....12 Varroa Mite……………………………………………………………………………....12 Tracheal Mite…………………………………………………………………………….25 Bee-Louse (Braula Coeca)………………………………………………...………..........28 Wax Moths (Achroia Grisella & Galleria Mellonella) ………………………………….29 Tropilaelaps Mite…………………………………………………...……………………31 Small Hive Beetle (SHB)……………………………………………………...…….……33 Rodents (Mice and Rats)…………………………………………………………………36 Wasps and Bumblebees…………………………………………………………………..37 Adult bee diseases……………………….………………………………………….................….38 Nosema…………………………………………………………………………………...38 Brood diseases.………………………………………...……………………….…………..….…40 American Foulbrood………………………………………………………..……………40 European Foulbrood………………………...…………………………………………...43 Chalkbrood ……………………………..…………………….………………………….45 Stonebrood ……………………………………………………………………………….47 Viral diseases…………………………………….…………………………….……...………….47 Sacbrood Virus (SBV)……...…………………………………………………………….47 Deformed Wing Virus (DWV)……………..…………………………………………………….49 Kashmir Virus (KBV)……………………………………………………………………………..50