Jennifer Szymanski Usfish and Wildlife Service Endangered
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
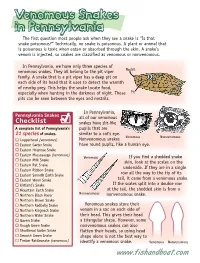
Venomous Snakes in Pennsylvania the First Question Most People Ask When They See a Snake Is “Is That Snake Poisonous?” Technically, No Snake Is Poisonous
Venomous Snakes in Pennsylvania The first question most people ask when they see a snake is “Is that snake poisonous?” Technically, no snake is poisonous. A plant or animal that is poisonous is toxic when eaten or absorbed through the skin. A snake’s venom is injected, so snakes are classified as venomous or nonvenomous. In Pennsylvania, we have only three species of venomous snakes. They all belong to the pit viper Nostril family. A snake that is a pit viper has a deep pit on each side of its head that it uses to detect the warmth of nearby prey. This helps the snake locate food, especially when hunting in the darkness of night. These Pit pits can be seen between the eyes and nostrils. In Pennsylvania, Pennsylvania Snakes all of our venomous Checklist snakes have slit-like A complete list of Pennsylvania’s pupils that are 21 species of snakes. similar to a cat’s eye. Venomous Nonvenomous Copperhead (venomous) Nonvenomous snakes Eastern Garter Snake have round pupils, like a human eye. Eastern Hognose Snake Eastern Massasauga (venomous) Venomous If you find a shedded snake Eastern Milk Snake skin, look at the scales on the Eastern Rat Snake underside. If they are in a single Eastern Ribbon Snake Eastern Smooth Earth Snake row all the way to the tip of its Eastern Worm Snake tail, it came from a venomous snake. Kirtland’s Snake If the scales split into a double row Mountain Earth Snake at the tail, the shedded skin is from a Northern Black Racer Nonvenomous nonvenomous snake. -

Ecology of the Desert Massasauga Rattlesnake in Colorado: Habitat and Resource Utilization
Ecology of the Desert Massasauga Rattlesnake in Colorado: Habitat and Resource Utilization A report to the CDOW and USFWS for the Colorado Wildlife Conservation Grant Program Dr. Stephen P. Mackessy School of Biological Sciences University of Northern Colorado 501 20th St., CB 92 Greeley, CO 80639-0017 USA Tel: (970)-351-2429 Fax: (970)-351-2335 Email: [email protected] http://www.unco.edu/nhs/biology/faculty_staff/mackessy_stephen.htm Desert Massasauga (Sistrurus catenatus edwardsii) from Lincoln County, Colorado ABSTRACT We conducted a radiotelemetric study on a population of Desert Massasaugas (Sistrurus catenatus edwardsii) in Lincoln County, SE Colorado. Massasaugas were most active between 14 and 30 °C, with an average ambient temperature during activity of 22.1°C. In the spring, snakes make long distance movements (up to 2 km) from the hibernaculum (shortgrass, compacted clay soils) to summer foraging areas (mixed grass/sandsage, sand hills). Summer activity is characterized by short distance movements, and snakes are most often observed at the base of sandsage in ambush or resting coils. Massasaugas gave birth to 5-7 young in late August, and reproduction appears to be biennial. Observations on three radioed gravid females indicated that Desert Massasaugas show maternal attendance for at least five days post-parturition. Snakes returned to the hibernaculum area in October and appear to hibernate individually in rodent burrows most commonly. However, the immediate region is utilized by several other species of snakes, particularly Prairie Rattlesnakes (Crotalus viridis viridis), and Massasaugas were also observed at the entrance to these den sites, suggesting that they are used by the Massasaugas as well. -

Agkistrodon Piscivorus)
BearWorks MSU Graduate Theses Fall 2019 Behavioral Aspects Of Chemoreception In Juvenile Cottonmouths (Agkistrodon Piscivorus) Chelsea E. Martin Missouri State University, [email protected] As with any intellectual project, the content and views expressed in this thesis may be considered objectionable by some readers. However, this student-scholar’s work has been judged to have academic value by the student’s thesis committee members trained in the discipline. The content and views expressed in this thesis are those of the student-scholar and are not endorsed by Missouri State University, its Graduate College, or its employees. Follow this and additional works at: https://bearworks.missouristate.edu/theses Part of the Behavior and Ethology Commons Recommended Citation Martin, Chelsea E., "Behavioral Aspects Of Chemoreception In Juvenile Cottonmouths (Agkistrodon Piscivorus)" (2019). MSU Graduate Theses. 3466. https://bearworks.missouristate.edu/theses/3466 This article or document was made available through BearWorks, the institutional repository of Missouri State University. The work contained in it may be protected by copyright and require permission of the copyright holder for reuse or redistribution. For more information, please contact [email protected]. BEHAVIORAL ASPECTS OF CHEMORECEPTION IN JUVENILE COTTONMOUTHS (AGKISTRODON PISCIVORUS) A Master’s Thesis Presented to The Graduate College of Missouri State University TEMPLATE In Partial Fulfillment Of the Requirements for the Degree Master of Science, Biology By Chelsea E. Martin December 2019 Copyright 2019 by Chelsea Elizabeth Martin ii BEHAVIORAL ASPECTS OF CHEMORECPTION IN JUVENILE COTTONMOUTHS (AGKISTRODON PISCIVORUS) Biology Missouri State University, December 2019 Master of Science Chelsea E. Martin ABSTRACT For snakes, chemical recognition of predators, prey, and conspecifics has important ecological consequences. -

The Eastern Massasauga Rattlesnake Is Toxic, but Only a Small Amount Is Injected Through Short Fangs
What’s so special about the massasauga? Northern Indiana’s The rare eastern massasauga is northern Indiana’s only native rattlesnake. Endangered Because eastern massasaugas are shy and secretive, you may never see one in the wild. These snakes hide under Rattlesnake brush and retreat to a sheltered area if spotted in the open. Although eastern massasaugas are venomous, they usually flee from humans rather than bite. Their venom Range of the Eastern Massasauga Rattlesnake is toxic, but only a small amount is injected through short fangs. The last human fatality from an eastern Why are they endangered? massasauga bite occurred more than 60 years ago. Eastern massasaugas are endangered over much of their Eastern massasaugas live in range because their wetland habitats are often drained and northern Indiana’s swamps, Eastern massasaugas play an important role in the filled for development. These snakes are also collected for bogs, and wetlands. ecosystem by feeding on mice, voles, and shrews, thus the illegal reptile trade. People may kill massasaugas out of Christopher Smith helping to keep the rodent population under control. fear, not realizing the snakes’ importance to the ecosystem. People and snakes can live together. Nicholas Scobel Understanding snakes is the first step. Learn to identify eastern massasaugas and other Indiana snakes. A field guide to native reptiles can help. Wear proper footwear when hiking in areas where snakes might be found, especially at night. Stay on trails. Limit the use of pesticides and other chemicals on natural areas on your property. All wildlife will benefit. Teach your family and friends about snakes and what to do if they see an eastern massasauga. -

Species Assessment for the Midget Faded Rattlesnake (Crotalus Viridis Concolor)
SPECIES ASSESSMENT FOR THE MIDGET FADED RATTLESNAKE (CROTALUS VIRIDIS CONCOLOR ) IN WYOMING prepared by 1 2 AMBER TRAVSKY AND DR. GARY P. BEAUVAIS 1 Real West Natural Resource Consulting, 1116 Albin Street, Laramie, WY 82072; (307) 742-3506 2 Director, Wyoming Natural Diversity Database, University of Wyoming, Dept. 3381, 1000 E. University Ave., Laramie, WY 82071; (307) 766-3023 prepared for United States Department of the Interior Bureau of Land Management Wyoming State Office Cheyenne, Wyoming October 2004 Travsky and Beauvais – Crotalus viridus concolor October 2004 Table of Contents INTRODUCTION ................................................................................................................................. 2 NATURAL HISTORY ........................................................................................................................... 2 Morphological Description........................................................................................................... 3 Taxonomy and Distribution ......................................................................................................... 4 Habitat Requirements ................................................................................................................. 6 General ............................................................................................................................................6 Area Requirements..........................................................................................................................7 -

Pit Vipers: from Fang to Needle—Three Critical Concepts for Clinicians
Tuesday, July 28, 2021 Pit Vipers: From Fang to Needle—Three Critical Concepts for Clinicians Keith J. Boesen, PharmD & Nicholas B. Hurst, M.D., MS Disclosures / Potential Conflicts of Interest • Keith Boesen and Nicholas Hurst are employed by Rare Disease Therapeutics, Inc. (RDT) • RDT is a U.S. company working with Laboratorios Silanes, S.A. de C.V., a company in Mexico • Laboratorios Silanes manufactures a variety of antivenoms Note: This program may contain the mention of suppliers, brands, products, services or drugs presented in a case study or comparative format using evidence-based research. Such examples are intended for educational and informational purposes and should not be perceived as an endorsement of any particular supplier, brand, product, service or drug. 2 Learning Objectives At the end of this session, participants should be able to: 1. Describe the venom variability in North American Pit Vipers 2. Evaluate the clinical symptoms associated with a North American Pit Viper envenomation 3. Develop a treatment plan for a North American Pit Viper envenomation 3 Audience Poll Question: #1 of 5 My level of expertise in treating Pit Viper Envenomation is… a. I wouldn’t know where to begin! b. I have seen a few cases… c. I know a thing or two because I’ve seen a thing or two d. I frequently treat these patients e. When it comes to Pit Viper envenomation, I am a Ssssuper Sssskilled Ssssnakebite Sssspecialist!!! 4 PIT VIPER ENVENOMATIONS PIT VIPERS Loreal Pits Movable Fangs 1. Russel 1983 -Photo provided by the Arizona Poison and Drug Information Center 1. -

Mating in Free-Ranging Neotropical Rattlesnakes, Crotalus Durissus: Is It Risky for Males?
Herpetology Notes, volume 14: 225-227 (2021) (published online on 01 February 2021) Mating in free-ranging Neotropical rattlesnakes, Crotalus durissus: Is it risky for males? Selma Maria Almeida-Santos1,*, Thiago Santos2, and Luis Miguel Lobo1 Field observations of the mating behaviour of snakes The male remained stretched out for about 20 minutes are scarce, probably because of the secretive nature and and showed no defensive posture even with the presence low encounter rates of many species (Sasa and Curtis, of the observer. We then noticed drops of blood on the 2006). In the Neotropical rattlesnake, Crotalus durissus vegetation and the hemipenis (Fig. 1 E-F). We could not Linnaeus, 1758, mating has been reported only in determine the origin of the blood, but we suggest two captive individuals (Almeida-Santos et al., 1999). Here nonexclusive hypotheses. The hemipenis spicules may we describe the first record of the mating behaviour of have hurt the female’s vagina while she was dragging the the Neotropical rattlesnake, Crotalus durissus, in nature male over a long distance. Alternatively, the male may (Fig. 1 A). have suffered an injury to the hemipenis while being Observations were made on 9 March 2017, at 14:54 h, dragged quickly by the female. The slow hemipenis a warm and sunny day (temperature = 27.1 oC; relative retraction and the male’s fatigue after copulation may humidity = 66%), in an ecotone between dry forest and better support the second hypothesis. Cerrado (Brazilian savannah) in Prudente de Morais, Potential costs for male C. durissus during mating Minas Gerais, Brazil (-19.2841 °S,-44.0628 °W; datum season include increased activity and energy expenditure WGS 84). -

Appendix I War of 1812 Chronology
THE WAR OF 1812 MAGAZINE ISSUE 26 December 2016 Appendix I War of 1812 Chronology Compiled by Ralph Eshelman and Donald Hickey Introduction This War of 1812 Chronology includes all the major events related to the conflict beginning with the 1797 Jay Treaty of amity, commerce, and navigation between the United Kingdom and the United States of America and ending with the United States, Weas and Kickapoos signing of a peace treaty at Fort Harrison, Indiana, June 4, 1816. While the chronology includes items such as treaties, embargos and political events, the focus is on military engagements, both land and sea. It is believed this chronology is the most holistic inventory of War of 1812 military engagements ever assembled into a chronological listing. Don Hickey, in his War of 1812 Chronology, comments that chronologies are marred by errors partly because they draw on faulty sources and because secondary and even primary sources are not always dependable.1 For example, opposing commanders might give different dates for a military action, and occasionally the same commander might even present conflicting data. Jerry Roberts in his book on the British raid on Essex, Connecticut, points out that in a copy of Captain Coot’s report in the Admiralty and Secretariat Papers the date given for the raid is off by one day.2 Similarly, during the bombardment of Fort McHenry a British bomb vessel's log entry date is off by one day.3 Hickey points out that reports compiled by officers at sea or in remote parts of the theaters of war seem to be especially prone to ambiguity and error. -

The Massasauga Rattlesnake
Pennsylvania Amphibians & Reptiles TTheh e MMassasauga s s a s a u g a R a t t l e s n a k e by Andrew L. Shiels “Massasauga,” “swamp rattler,” and “black snapper,” are a 24 to 30 inches. It has a grayish body color with a series of few of the nicknames given to a rare, secretive, yet interest- dark-brown, irregularly shaped blotches along the top of ing rattlesnake found in northwestern Pennsylvania. Its the back. These blotches are the source of the Latin spe- limited distribution and unique ecological needs have un- cies name “catenatus,” which means “chain-like.” Along fortunately placed this species among Pennsylvania’s most the sides of the body are two to three rows of smaller, more endangered. Out of sight and out of mind to most Penn- rounded spots. sylvanians, the massasauga has been quietly disappearing Massasaugas are pit vipers, which means there is a for years. However, recent conservation efforts have been heat-sensing pit on the side of the head between the eye undertaken to ensure its survival in the Commonwealth. and nostril. Like our other two venomous species, the Understanding its biology and ecology are keys to devel- northern copperhead and the eastern timber rattlesnake, oping an effective conservation strategy. massasaugas have vertically elliptical pupils and a single The eastern massasauga (Sistrurus catenatus catenatus) row of scales on the underside of the tail. Unlike timber is a medium-sized venomous snake that reaches lengths of rattlesnakes, which have many small scales on the head, a massasauga’s head is covered with nine large, scale plates typical of our non-venomous species. -

Eastern Diamondback Rattlesnake (Crotalus Adamanteus) Ambush Site Selection in Coastal Saltwater Marshes
Marshall University Marshall Digital Scholar Theses, Dissertations and Capstones 2020 Eastern Diamondback Rattlesnake (Crotalus adamanteus) Ambush Site Selection in Coastal Saltwater Marshes Emily Rebecca Mausteller [email protected] Follow this and additional works at: https://mds.marshall.edu/etd Part of the Aquaculture and Fisheries Commons, Behavior and Ethology Commons, Other Ecology and Evolutionary Biology Commons, and the Terrestrial and Aquatic Ecology Commons Recommended Citation Mausteller, Emily Rebecca, "Eastern Diamondback Rattlesnake (Crotalus adamanteus) Ambush Site Selection in Coastal Saltwater Marshes" (2020). Theses, Dissertations and Capstones. 1313. https://mds.marshall.edu/etd/1313 This Thesis is brought to you for free and open access by Marshall Digital Scholar. It has been accepted for inclusion in Theses, Dissertations and Capstones by an authorized administrator of Marshall Digital Scholar. For more information, please contact [email protected], [email protected]. EASTERN DIAMONDBACK RATTLESNAKE (CROTALUS ADAMANTEUS) AMBUSH SITE SELECTION IN COASTAL SALTWATER MARSHES A thesis submitted to the Graduate College of Marshall University In partial fulfillment of the requirements for the degree of Master of Science In Biological Sciences by Emily Rebecca Mausteller Approved by Dr. Shane Welch, Committee Chairperson Dr. Jayme Waldron Dr. Anne Axel Marshall University December 2020 i APPROVAL OF THESIS We, the faculty supervising the work of Emily Mausteller, affirm that the thesis, Eastern Diamondback Rattlesnake (Crotalus adamanteus) Ambush Site Selection in Coastal Saltwater Marshes, meets the high academic standards for original scholarship and creative work established by the Biological Sciences Program and the College of Science. This work also conforms to the editorial standards of our discipline and the Graduate College of Marshall University. -

Lesser Known Snakes (PDF)
To help locate food, snakes repeat- edly flick out their narrow forked tongues to bring odors to a special sense organ in their mouths. Snakes are carnivores, eating a variety of items, including worms, with long flexible bodies, snakes insects, frogs, salamanders, fish, are unique in appearance and small birds and mammals, and even easily recognized by everyone. other snakes. Prey is swallowed Unfortunately, many people are whole. Many people value snakes afraid of snakes. But snakes are for their ability to kill rodent and fascinating and beautiful creatures. insect pests. Although a few snakes Their ability to move quickly and are venomous, most snakes are quietly across land and water by completely harmless to humans. undulating their scale-covered Most of the snakes described bodies is impressive to watch. here are seldom encountered— Rather than moveable eyelids, some because they are rare, but snakes have clear scales covering most because they are very secre- their eyes, which means their eyes tive, usually hiding under rocks, are always open. Some snake logs or vegetation. For information species have smooth scales, while on the rest of New York’s 17 species others have a ridge on each scale of snakes, see the August 2001 that gives it a file-like texture. issue of Conservationist. Text by Alvin Breisch and Richard C. Bothner, Maps by John W. Ozard (based on NY Amphibian & Reptile Atlas Project) Artwork by Jean Gawalt, Design by Frank Herec This project was funded in part by Return A Gift To Wildlife tax check-off, U.S. Fish & Wildlife Service and the NYS Biodiversity Research Institute. -

Detroit Heritage River Nomination Report
Nomination of the Detroit River Submission Requirements This document sets out the criteria of the Canadian Heritage Rivers Board (CHRS) for the nomination of the Detroit River as a Heritage River within the Canadian Heritage Rivers System (CHRS). The CHRS is the vehicle by which the nominating agency identifies the heritage significance of a river and justifies its inclusion in the CHRS. The purpose of this format is to act as a guide to nominating agencies and to provide consistency in the nomination process. CHRS provides an opportunity for the recognition and conservation of rivers deemed to be of outstanding Canadian heritage value. This value is obtained when it has been determined that a river is an outstanding representative of or unique in a province or territory. By the inclusion of such rivers in a single coast-to-coast system, they become representative of Canada’s river heritage as a whole, thus reflecting a “Canadian value”. 1 Nomination of the Detroit River Canadian Heritage Rivers System The Canadian Heritage Rivers System (CHRS) has been established by the federal, provincial and territorial governments to recognize outstanding rivers of Canada and ensuring management which will protect these rivers and enhance their significant heritage values for the long term benefit and enjoyment of Canadians. To qualify for the Canadian Heritage Rivers System, a river or section of a river must be of outstanding significance in one or more areas: natural heritage, human heritage, or recreational values. The nominated section should be large enough to encompass these values and provide the user with an appreciation of the river’s resources, as well as an enjoyable recreational experience.