An Interpretation of Oncoid Mass-Occurrence During the Late Silurian Lau Event, Gotland, Sweden
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

VA-Utbyggnadsplan För Region Gotland Plan För En Hållbar Utbyggnad Av VA-Försörjningen
VA-plan 2018 VA-utbyggnadsplan för Region Gotland Plan för en hållbar utbyggnad av VA-försörjningen Möjligheten att försörja nuvarande och tillkommande VA-planeringens olika dokument behöver hållas aktuella invånare och besökare med vatten har under de senaste för att de ska vara ett användbart underlag i regionens åren varit en begränsande faktor för exploatering och arbete med VA-frågor och översiktliga planering. Allt utveckling av bebyggda områden på flera platser på Got- eftersom VA-utbyggnadsplanens åtgärder genomförs land. Vision och strategi för Gotlands VA-försörjning 2030 skapas ett nytt nuläge som blir utgångs¬punkt för över- som antogs av Regionfullmäktige den 18 december 2017 syn av VA-strategin. En avstämning av VA-utbyggnads- är tydlig. Regionens arbete med VA-försörjning behöver planens åtgärder och införlivning av åtgärderna i verk- ske med utgångspunkt i tillgången till tjänligt dricksvat- samhetens budgetplanering sker årligen. ten och möjligheten att ordna en godkänd avloppsan- läggning – på hela ön. Region Gotland tar ett helhetsgrepp kring långsiktigt hållbar planering av dricksvatten, spillvatten och dag- vatten. Arbetet sker stegvis. Detta dokument är en VA-ut- byggnadsplan för Region Gotland som tillsammans med andra delplaner skapar en samlad VA-plan. Ansvaret för VA-utbyggnadsplanen ligger hos Tekniska förvaltningen, därtill berörs flera andra förvaltningar av planen. ÖNSKAT LÄGE 2030 • En beskrivning av ett fram- tida läge för VA-försörjningen på Gotland som ger alla berörda VA-PLAN samma syn på vart investeringar och arbete ska leda. • Sammanställning av åtgärder som behövs för Antagen av regionfullmäktige att i enlighet med strategin, 2017-12-18 STRATEGI gå från nuläget till ett önskat • Tydliga riktlinjer om vad som läge (målbild/vision). -

Reference Localities for Palaeontology and Geology in the Silurian of Gotland
SVERIGES GEOLOGISKA UNDERSOKNING SER C NR 705 AVHANDLINGAR OCH UPPSATSER ÅRSBOK 68 NR 12 SVEN LAUFELD REFERENCE LOCALITIES FOR PALAEONTOLOGY AND GEOLOGY IN THE SILURIAN OF GOTLAND STOCKHOLM 1974 SVERIGES GEOLOGISKA UNDERSOKNING SER C NR 705 AVHANDLINGAR OCH UPPSATSER ÅRSBOK 68 NR 12 SVEN LAUFELD REFERENCE LOCALITIES FOR PALAEONTOLOGY AND GEOLOGY IN THE SILURIAN OF GOTLAND STOCKHOLM 1974 ISBN 91-7158-059-X Kartorna är godkända från sekretessynpunkt för spridning Rikets allmänna kartverk 1974-03-29 IU S UNES D l Project ECOSTRATIGRAPHY Laufeld, S.: Reference loca!ities for palaeontology and geology in the Silurian of Gotland. Sveriges Geologiska Undersökning, Ser. C, No. 705, pp. 1-172. Stock holm, 24th May, 1974. About 530 geologkal localities in the Silurian of the island of Gotland, Sweden, are described under code names in alphabetical order. Each locality is provided with a UTM grid reference and a detailed description with references to the topographical and geologkal map sheets. Information on reference points and levels are included for some localities. The stratigraphic position of each locality is stated. A bibliography is attached to several localities. Sven Laufeld, Department of Historical Geology and Palaeontology, Sölvegatan 13, S-223 62 Lund, Sweden, 4th March, 1974. 4 Contents Preface. By Anders Martinsson 5 Introduction 7 Directions for use lO Grid references 10 Churches 11 Detailed descriptions 11 Reference point and leve! 11 Stratigraphy 11 References 12 Indexes .. 12 Practical details 13 Descriptions of localities 14 References .. 145 Index by topographical maps 149 Index by geological maps 157 Index by stratigraphical order .. 165 5 Preface In 1968 a course was set for continued investigations of the Silurian of Gotland and Scania. -

(Foram in Ifers, Algae) and Stratigraphy, Carboniferous
MicropaIeontoIogicaI Zonation (Foramin ifers, Algae) and Stratigraphy, Carboniferous Peratrovich Formation, Southeastern Alaska By BERNARD L. MAMET, SYLVIE PINARD, and AUGUSTUS K. ARMSTRONG U.S. GEOLOGICAL SURVEY BULLETIN 2031 U.S. DEPARTMENT OF THE INTERIOR BRUCE BABBITT, Secretary U.S. GEOLOGICAL SURVEY Robert M. Hirsch, Acting Director Any use of trade, product, or firm names in this publication is for descriptive purposes only and does not imply endorsement by the U.S. Government Text and illustrations edited by Mary Lou Callas Line drawings prepared by B.L. Mamet and Stephen Scott Layout and design by Lisa Baserga UNITED STATES GOVERNMENT PRINTING OFFICE, WASHINGTON : 1993 For sale by Book and Open-File Report Sales U.S. Geological Survey Federal Center, Box 25286 Denver, CO 80225 Library of Congress Cataloging in Publication Data Mamet, Bernard L. Micropaleontological zonation (foraminifers, algae) and stratigraphy, Carboniferous Peratrovich Formation, southeastern Alaska / by Bernard L. Mamet, Sylvie Pinard, and Augustus K. Armstrong. p. cm.-(U.S. Geological Survey bulletin ; 2031) Includes bibtiographical references. 1. Geology, Stratigraphic-Carboniferous. 2. Geology-Alaska-Prince of Wales Island. 3. Foraminifera, Fossil-Alaska-Prince of Wales Island. 4. Algae, Fossil-Alaska-Prince of Wales Island. 5. Paleontology- Carboniferous. 6. Paleontology-Alaska-Prince of Wales Island. I. Pinard, Sylvie. II. Armstrong, Augustus K. Ill. Title. IV. Series. QE75.B9 no. 2031 [QE671I 557.3 s--dc20 [551.7'5'097982] 92-32905 CIP CONTENTS Abstract -

ABSTRACTS BOOK Proof 03
1st – 15th December ! 1st International Meeting of Early-stage Researchers in Paleontology / XIV Encuentro de Jóvenes Investigadores en Paleontología st (1December IMERP 1-stXIV-15th EJIP), 2018 BOOK OF ABSTRACTS Palaeontology in the virtual era 4 1st – 15th December ! Ist Palaeontological Virtual Congress. Book of abstracts. Palaeontology in a virtual era. From an original idea of Vicente D. Crespo. Published by Vicente D. Crespo, Esther Manzanares, Rafael Marquina-Blasco, Maite Suñer, José Luis Herráiz, Arturo Gamonal, Fernando Antonio M. Arnal, Humberto G. Ferrón, Francesc Gascó and Carlos Martínez-Pérez. Layout: Maite Suñer. Conference logo: Hugo Salais. ISBN: 978-84-09-07386-3 5 1st – 15th December ! Palaeontology in the virtual era BOOK OF ABSTRACTS 6 4 PRESENTATION The 1st Palaeontological Virtual Congress (1st PVC) is just the natural consequence of the evolution of our surrounding world, with the emergence of new technologies that allow a wide range of communication possibilities. Within this context, the 1st PVC represents the frst attempt in palaeontology to take advantage of these new possibilites being the frst international palaeontology congress developed in a virtual environment. This online congress is pioneer in palaeontology, offering an exclusively virtual-developed environment to researchers all around the globe. The simplicity of this new format, giving international projection to the palaeontological research carried out by groups with limited economic resources (expensive registration fees, travel, accomodation and maintenance expenses), is one of our main achievements. This new format combines the benefts of traditional meetings (i.e., providing a forum for discussion, including guest lectures, feld trips or the production of an abstract book) with the advantages of the online platforms, which allow to reach a high number of researchers along the world, promoting the participation of palaeontologists from developing countries. -

Microscopic Microbialite Textures and Their Components in a Lower Devonian Lagoonal Facies of the Fukuji Formation, Gifu Prefecture, Central Japan
Microscopic microbialite textures and their components in a Lower Devonian lagoonal facies of the Fukuji Formation, Gifu Prefecture, central Japan Natsuko ADACHI, Yoichi EZAKI ADACHI, N. & EZAKI, Y., 2007: Microscopic microbialite textures and their components in a Lower Devo- nian lagoonal facies of the Fukuji Formation, Gifu Prefecture, central Japan. – In: HUBMANN, B. & PILLER, W. E. (Eds.): Fossil Corals and Sponges. Proceedings of the 9th International Symposium on Fossil Cni- daria and Porifera. – Österr. Akad. Wiss., Schriftenr. Erdwiss. Komm. 17: 215–229, 2 Figs., 2 Pls., Wien. Abstract: Varied microbialite textures together with skeletal metazoans such as tabulate and rugose corals and stromatoporoids are present in the Lower Devonian lagoonal Fukuji Formation, Gifu Prefecture, central Japan. Stromatolitic, thrombolitic, and leiolitic textures that have formed around the skeletons comprise three main components: peloids, micrites, and calcimicrobes. The calcimi- crobe Girvanella is ubiquitous and is closely associated with the formation of varied microbialite (stromatolitic, thrombolitic, and leiolitic) textures as accumulations, partial aggregations, and ran- dom distributions. Peloids and micrites also exhibit varied microbialite textures by manners of assemblage similar to those in Girvanella. Girvanella itself in some cases gives rise to peloids and micrites. In contrast, Rothpletzella and Renalcis relate only to the formation of particular micro- bialite textures such as stromatolitic and thrombolitic textures, respectively. Microbialite textures have not yet been fully examined in Devonian lagoonal environments. However, it is emphasized that Girvanella was comparatively abundant there and played an important role in the genesis of varied microbialite textures and their components, as seen in the lagoonal Fukuji Formation. Key words: Microbialite textures, components, Girvanella, lagoonal environment, Lower Devo- nian, Japan Contents 1. -

Adress Socken/Tätort Ada Blocks Gata Visby Adelsgatan Visby Innerstad
Adress Socken/Tätort Ada Blocks gata Visby Adelsgatan Visby Innerstad Adelsgatan Visby Innerstad Ainbuskstigen Sproge Akarvägen Sproge Aklejastigen Silte Algatan Romakloster Alkviar Hellvi Allégatan Visby Allhagevägen Västerhejde Almedalen Visby Innerstad Almvägen Visby Alvstigen Tofta Anders Wedins väg Västerhejde Andersrogatan Visby Andrégatan Visby Anemonvägen Visby Angelbosvägen Lärbro Aniaragatan Visby Anna Dubbes väg Katthammarsvik Annexgatan Romakloster Apelgatan Visby Apgränd Visby Innerstad Apoteksgatan Hemse Apoteksgatan Slite Appellstigen Visby Aprilvägen Åminne Arkosvägen Stenkyrka Artilleribacken Visby Innerstad Artillerigatan Visby Artilleriparken Visby Artillerivägen Fårösund Asarvegatan Hemse Asbrovägen Åminne Aspgatan Visby Asplunds väg Vibble Atlasgatan Visby Audungsvägen Fårösund Augustivägen Åminne Aurvägen Fårösund Avagatan Visby Axel Ekmans väg Västerhejde Axstigen Silte Backérsvägen Klintehamn 2016-11-22 Backgatan Visby Backskogsvägen Västerhejde Backvägen Fårösund Badhusgatan Slite Badvägen Ljugarn Balders väg Fröjel Baldersgatan Visby Bandavägen Tofta Bandystigen Slite Bangårdsgatan Hemse Bangårdsgatan Visby Banvallsvägen Klintehamn Barlastvägen Klintehamn Barrgränd Lärbro Barrvägen Eskelhem Basgången Vibble Beckatorpsvägen Västerhejde Bendix park Klintehamn Bergbetningen Visby Bergbetningen Visby Berggränd Visby Innerstad Bergmansgatan Visby Bergsgatan Visby Bergsvägen Väskinde Betesgatan Visby Bingebygatan Visby Bingebyparken Visby Biotitvägen Stenkyrka Birgers gränd Visby Innerstad Birkagatan Visby Biskopsgatan -

Iberopora Bodeuri GRANIER & BERTHOU 2002 (Incertae Sedis)
Geologia Croatica 57/1 1–13 2 Figs. 2 Pls. ZAGREB 2004 Iberopora bodeuri GRANIER & BERTHOU 2002 (incertae sedis) from the Plassen Formation (Kimmeridgian–Berriasian) of the Tethyan Realm Felix SCHLAGINTWEIT Key words: Incertae sedis, Upper Jurassic, Lower contribution, the new microproblematicum Iberopora n. Cretaceous, Plassen Formation, Northern Calcareous gen. with the type-species Iberopora bodeuri n.sp. was Alps, Austria. reported from the Berriasian of Portugal by GRANIER & BERTHOU (2002). An adequate description (without generic, species diagnosis) was presented, but little Abstract information was provided concerning other occurrences, Iberopora bodeuri GRANIER & BERTHOU 2002, formerly known or facies, and stratigraphic data. As only one reference as “crust problematicum” (SCHMID, 1996) is described from the Plassen Formation (Kimmeridgian–Berriasian) of the Northern (the type region) was cited in the synonymy, almost Calcareous Alps (NCA). Here, it occurs either as an incrustation on nothing is known about its palaeogeographic distribu- corals/stromatoporoids or it forms nodular masses (“solenoporoid tion. In this discussion regarding Upper Jurassic to morphotype”). It is typically found in the fore-reef facies of the Lower Cretaceous material from the Northern Calca- platform margin, and (upper) slope deposits where autochthonous dasycladales are absent. Water turbulence appears to control the reous Alps of Austria and already published in part morphological development of Iberopora. Thus, flat crusts appear in (SCHLAGINTWEIT & EBLI, 1999) the species was less agitated settings. The crusts are almost always accompanied by refered to as “Krustenproblematikum” SCHMID, calcareous sponges/sclerosponges and abundant micro-encrusters, mostly Koskinobullina socialis CHERCHI & SCHROEDER and 1996). Here we summarize the available data to provide “Tubiphytes” morronensis CRESCENTI. -

Inventering Av Naturvärden I Marina Kustområden
Inventering av naturvärden i marina kustområden Rapporter om natur och miljö – nr 2007: 14 2 Inventering av naturvärden i marina kustområden Underlag inför bildande av eventuella marina reservat Magnus Petersson Castor & Pollux Castor & Pollux bedriver sin huvudsakliga verksamhet inom området för akvatisk konsultation. Företaget innehar betydande erfarenhet av miljöövervakningsarbete. Företaget uppvisar även bred kompetens inom området för undervattensdokumentation i form av dykning, fotografering och videofilmning. Kontakta Castor & Pollux genom att ringa Magnus Petersson, 0737-165110 Omslagsbild: Omslagsfotografier tagna av Magnus Petersson. ISSN 1653—7041 LÄNSSTYRELSEN I GOTLANDS LÄN – VISBY 2007 3 Förord Denna rapport har upprättats på uppdrag av Länsstyrelsen i Gotlands län. Syftet med undersökningen var att inventera naturvärden i två områden längs Gotlands östra kust inför eventuellt bildande av marina reservat. Ansvarsförhållanden För innehållet i denna rapport ansvarar författaren Magnus Petersson. Fotografier/bilder Samtliga fotografier är tagna av Magnus Petersson. Fotografierna i föreliggande rapport är skyddade enligt lagen om upphovsrätt och tillhör fotografen. Fotografen ger sitt medgivande till intern (Länsstyrelsen Gotland) användning av fotografierna vid t ex presentationer. Mångfaldigande av fotografierna i form av tryckning, kopiering etc är förbjudet. Kontaktperson på Länsstyrelsen Peter Landergren, Vatten och fiske, Länsstyrelsen i Gotlands län, 621 85 Visby, tel 0498/292119. 4 Innehållsförteckning Sammanfattning -
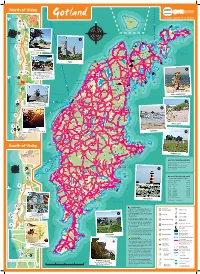
Gotlandscykeluthyrning.Com, +46 498 214 133 Lumme- Coustline up Gotland Lunda to Fårö Appr
North of Visby Take the Fyrplatsen gotlandscykeluthyrning.com, +46 498 214 133 Lumme- coustline up Gotland lunda to Fårö appr. 15 km 70 km from Gotska Sandön from Visby. N Kyrkudden Visby Östersjön Gotska Sandön Langhammars 2 Norsholmen Hamnudden Tärnudden Tälle- N Helgumannen vika Ekeviken Ekeviken Skär 149 Norsta Auren 14 Digerhuvud Butleks Avanäs 3 Kalbjärga 14 Norrsund Ullahau Fårö fyr Lauterhorn Lauter Ava Sudersand Sudersandsviken Blå Gamlehamn 1 Lagunen Hallshuk Norra gattet Mölnor Ar Lansa FÅRÖ Fårö infocenter Brissund Fårö kyrka Kappelshamns- viken Hammars Bläse Kalkbruksmuseum Bästeträsk Häftingsklint Northen coastline HALL Fårösund Dämba FLERINGE Östersjön Kappels- FÅRÖSUND Sigsarve hamn strand Bräntings haid Badhus- Engelska BUNGE Kolerakyrkogården Irevik Bungemuseet parken 13 Hideviken KAPPELS- Ryssnäs HAMN 148 Gotland Ring Jungfrun Ireviken Lickershamn HANGVAR Visby RUTE Stenkyrkehuk Gamla Bungeviken Bungeör 149 Thor Bungenäs 149 Fardume träsk Fardume slott Skenholmen Fardume Lergrav Hångers källa Valleviken 2 STENKYRKA träsk LÄRBRO Själsö Bageri – Bakery & Café Valle- vik Garden café with beatiful sea views. Nyhamn LUMMELUNDA HELLVI Husken Furillen Enjoy tartin and a glass of wine at sunset. Kyllaj Själsö Open from 5 pm. Welcome! 148 13 Brissund Kyllaj Brissundsvägen 22A, +46 498 270 909 MARTEBO 8 km OTHEM 147 Lummelundagrottan Hide from Själsö Quick TINGSTÄDE Visby Seaside bikepath Tingstäde- File haidar Hideviken Krusmyntagården träsk Vägume- bikepath viken Hide- Brissund viken 148 S:t Olofsholm 12 Nynäshamn Hejnum hällar SLITE Asunden 149 Slite havsbad Själsö Boge- Kallgateburg vik Enholmen 149 Brucebo VÄSKINDE LOKRUME HEJNUM BOGE Snäck Östersjön BRO Bogeklinten Rauk, Langhammars Gustafsvik 147 Norderstrand FOLE 146 1 148 BÄL Tjälderviken Västervik 3 The Snäck Chimpansee rauk 147 HEJDEBY Tjelvars grav Tjälder 12 The Closest rauk from Visby appr. -

(Brachiopoda, Strophomenida) from the Silurian of Gotland, Sweden
Pala¨ontol Z (2011) 85:201–229 DOI 10.1007/s12542-010-0088-3 RESEARCH PAPER Strophomenidae, Leptostrophiidae, Strophodontidae and Shaleriidae (Brachiopoda, Strophomenida) from the Silurian of Gotland, Sweden Ole A. Hoel Received: 23 September 2010 / Accepted: 1 October 2010 / Published online: 13 November 2010 Ó The Author(s) 2010. This article is published with open access at Springerlink.com Abstract Twelve species of Brachiopods are described short ranged and occurs in low-energy environments in the from the Silurian of Gotland, six furcitellinines and six latest Llandovery. The species belonging to the Stroph- ‘‘strophodontids.’’ One is new—Strophodonta hoburgensis odontidae (Strophodonta hoburgensis n. sp.) and Shalerii- n. sp. The furcitellinines are moderately common and dae [Shaleria (Janiomya) ornatella and S. (Shaleriella) diverse in the lower part of the succession, but the last ezerensis] occur only in high-energy environments and have species disappears in the middle Hemse beds (*middle a short range within the late Ludlow. Ludlow). Three genera are represented: Bellimurina, Pentlandina and Katastrophomena, with the species and Keywords Silurian Á Llandovery Á Wenlock Á Ludlow Á subspecies B. wisgoriensis, P. tartana, P. loveni, P. lewisii Strophomenide Á Furcitellininae Á Leptostrophiidae Á lewisii, K. penkillensis and K. antiquata scabrosa. Most of Strophodontidae Á Shaleriidae the taxa are confined to low energy environments, but P. loveni was evidently specialized for the high energy reef Kurzfassung Zwo¨lf arten Brachiopoden von der Silurium environments of the Ho¨gklint Formation. B. wisgoriensis Gotlands sind beschrieben. Sechs Furcitellininen und sechs displays environmentally induced morphological variability ,,Strophodonten‘‘. Eine Gattung ist neu; Strophodonta in developing strong, frilly growth lamellae in high-energy hoburgensis sp. -

FACIES and Palaeoecology of the UPPER MEMBER of the AISEMONT FORMATION (Late Frasnian, S
GEOLOGICA BELGICA (2010) 13/3: 197-212 FACIES AND PALAEOECOLOGY OF THE UPPER MEMBER OF THE AISEMONT FORMATION (Late FRASNIAN, S. BELGIUM): AN UNUSUAL EPISODE WITHIN THE LATE FRASNIAN CRISIS Julien DENAYER & Edouard POTY (12 figures) Service de Paléontologie animale et humaine, Département de Géologie, Université de Liège. Bâtiment B18, Sart Tilman, 4000 Liège, Belgium. Email: [email protected]; [email protected] ABSTRACT The upper member of the Aisemont Formation – also known as the “second biostrome” - is the last significant Upper Frasnian carbonate unit in the northern part of the Namur-Dinant Basin (Southern Belgium). It consists of bioclastic limestone, often dolomitized, with numerous oncoids and corals. Despite its local name, the member is not a biostrome because only one thin bed is constructed by corals. It is mainly composed of limestone with numerous oncoids and a rich fauna of opportunistic organisms (bryozoans, brachiopods, gastropods, sponges, etc.) where Phillipsastrea and Frechastraea are the dominant coral taxa, associated with Alveolites. Ragged colonies of corals, as well as multi-encrusted bodies, show that the sea floor was soft and the rate of sedimentation was seasonal. These factors were unfavourable to common reef builders (stromatoporoids), thus the occurrence of one bed constructed by corals is a remarkable event that corresponds to the colonization of a hard ground defining the base of a falling stage systems tract. This confirms the model of the Aisemont sequence (third-order transgression-regression cycle). The abundant development of microbial structures in the member (oncoids) and in its lateral equivalent in stromatolites and thrombolites of the Petit-Mont Member (Les Valisettes Formation) in the Philippeville Anticlinorium is interpreted as an evidence of the environmental deterioration corresponding to the Late Frasnian Crisis. -

Abhandlungen Der Bayerischen Akademie Der Wissenschaften
© Biodiversity Heritage Library, http://www.biodiversitylibrary.org/; www.biologiezentrum.at Abhandlungen der Bayerischen Akademie der Wissenschaften Mathematisch - physikalische Klasse XXIX. Band, 5. Abhandlung Über die systematische Deutung und die stratigraphische Stellung der ältesten Versteinerungen Europas und Nordamerikas mit besonderer Berücksichtigung der Oryptozoen und Oolithe. III. T Über Oolithe. Nachgelassenes Manuskript von August Rothpletz, ergänzt durch eine von Rothpletz testamentarisch veranlaßte biologische Untersuchung rezenter Oolithe von Karl Giesenhagen. Mit 1 Tafel und 12 Textfiguren. Vorgelegt am 3. Dezember 1921 München 1922 Verlag der Bayerischen Akademie der Wissenschaften in Kommission des 6. Franzsehen Verlags (J. Roth) © Biodiversity Heritage Library, http://www.biodiversitylibrary.org/; www.biologiezentrum.at © Biodiversity Heritage Library, http://www.biodiversitylibrary.org/; www.biologiezentrum.at Einleitung zu der Arbeit von Giesenhagen über die Oolithe von Meskoutine. (Geschrieben von A. Rothpletz, Januar 1918.) Das Oolithen-Problem. Oolithe haben sich zu allen Zeiten vom Praekambrium an bis in die Gegenwart in erstaunlichen Mengen gebildet. Ihre eigenartige Form reizte schon im Altertum die Natur- forscher zu Erklärungen, für die wir heute meist nicht viel mehr als ein überlegenes Lächeln übrig haben. Doch darf man uns nur mit der Frage überraschen, was wir denn jetzt eigentlich über die Entstehung der Oolithen wissen, um jene Überlegenheit sofort in Verlegenheit umzuwandeln, und wenn es uns vielleicht gelingt, die Neugierde der Frager durch Mitteilung einer Fülle von neuen Beobachtungen und Vermutungen zu befriedigen, so raunt uns doch unser Inneres zu, daß wir über diesen Gegenstand noch heute eine be- stimmte Antwort zu geben nicht imstande sind. Noch niemanden ist es geglückt, das Wachsen eines Oolithen in der Natur zu beobachten oder im Laboratorium auf künstliche Weise einen echten Oolithen zu erzeugen.