Audition and Hemispheric Specialization in Songbirds and New Evidence from Australian Magpies
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Mount Canobolas Fauna Survey
MOUNT CANOBOLAS FAUNA SURVEY 24TH – 29th MARCH 2019 1 Introduction Mount Canobolas State Conservation Area The Mount Canobolas State Conservation Area (SCA) Draft Plan of Management begins with the following vision for this reserve: Mount Canobolas State Conservation Area is a sentinel in the landscape. It is a park valued, understood and protected by the community for its uniqueness, biodiversity, visitor experiences, and for the spiritual and cultural connections between people and this special place, now and into the future (OEH 2018). This iconic place is an extinct volcano rising to an altitude of 1397m asl and more than 500m above the surrounding central tablelands landscape. It has great significance for the local Wiradjuri custodians. The ecological values of this place are shaped by its high altitude, volcanic origins, isolation from similar geological formations and the extensive clearing of the surrounding landscape leaving it as a relatively isolated remnant of native vegetation. The presence of sub-alpine communities is particularly noteworthy (OEH 2018). The SCA incorporates an area of 1672 ha. The vegetation communities of the Mt Canobolas SCA have been broadly described and mapped with the following communities defined: 1. Eucalyptus macrorhyncha – E. dives: shrubby open forest and woodland (26% of the SCA) a. E. macrorhyncha dominant in more exposed sites b. E. dives dominant in less exposed sites 2. E. pauciflora – E. dalrympleana: Grassy woodlands and tall open forests (51% of the SCA) a. E. pauciflora – E. canobolensis grassy woodlands b. E. dalrympleana – E. viminalis tall open forests 3. Outcrop low open woodland (E. canobolensis – E. petroc?) and / or shrubs 4. -

Report No. 201601Rp1(F) AMBROSE
Report No. 201601rp1(f) AMBROSE ECOLOGICALSERVICES PTY. LTD (ABN: 34 097 016 496. ACN: 097 016 496) CLIENT ngh Environmental Pty Ltd, PO Box 470, Bega NSW 2550 Contact: Mr Nick Graham-Higgs, Mobile: 0427 260 819 Email: [email protected] This report has been prepared in accordance with the scope of services described in the contract or agreement between Ambrose Ecological Services Pty Ltd (Ambecol) and the Client. The report relies upon data, surveys, measurements and results taken at or under the particular times and conditions specified herein. Any findings, conclusions or recommendations only apply to the aforementioned circumstances and no greater reliance should be assumed or drawn by the Client. The products and services referred to in the report should not be regarded as Ambecol’s endorsement of specific manufacturers, distributors or brands. Furthermore, the report has been prepared solely for use by the Client and Ambecol accepts no responsibility for its use by other parties. Author: Dr Stephen Ambrose Position: Director, Ambrose Ecological Services Pty Ltd Signed: Date: 9 September 2016 2______________________________________AMBROSE ECOLOGICAL SERVICES PTY LTD Contents Sections Page No. 1 Introduction 5 2 Proposed Development 5 2.1 Subject Site 5 2.2 Proposed Turbine Development 10 3 Bird Survey and Assessment Methods 10 3.1 Overview 10 3.3 Existing Records 10 3.3 Field Surveys 14 4 Results 15 4.1 Overview 15 4.2 Field Studies 15 4.3 Threatened Bird Taxa 17 4.4 Migratory Bird Taxa 18 4.5 Island Endemics 19 5 Potential Impacts 52 5.1 Introduction 52 5.2 Analysis of Turbine Options for Lord Howe Island 52 5.3 Bird Impacts of Wind Turbines in Other Studies 59 5.4 Impacts on Threatened Bird Species 61 5.5 Impacts on Nationally-listed Migratory Bird Species 62 6 Recommendations 62 7 References 64 Appendices A Flesh-footed Shearwater Report ( & Carlile, 2015) 67 B Directions of Bird Flights Over Subject Site, February and March 77 2016 C Bird Flights Over Each Survey Area on Subject Site, February and 81 March 2016. -

Additional Records of Passerine Terrestrial Gaits
ADDITIONAL RECORDS OF PASSERINE TERRESTRIAL GAITS GEORGE A. CLARK, JR. The varied methods of locomotion in birds pose significant problems in behavior, ecology, adaptation, and evolution. On the ground birds progress with their legs moving either synchronously (hopping) or asynchronously (walking, running) as the extreme conditions. Relatively terrestrial species often have asynchronous gaits, whereas primarily arboreal species are typically synchronous on the ground. Particularly important earlier studies on passerines are Kunkels’ (1962) comparative behavioral survey and Riiggebergs’ (1960) analysis of the morphological correlates of gaits. Over several years I have noted gaits for 47 passerine species in the U.S., En- gland, and Kenya, and have examined many references. I here sum- marize behavioral records for families not mentioned by Kunkel (1962) and also for species with gaits markedly unlike those of confamilial species discussed by him. My supplementary review is selective rather than ex- haustive with the aim of indicating more fully the distribution of gaits among the passerine families. Regional handbooks, life history studies, and other publications contain numerous additional records, but I know of none that negate the conclusions presented here. J. S. Greenlaw (in prep.) has reviewed elsewhere the passerine double-scratch foraging be- havior that has at times previously been discussed in connection with gaits (e.g., in Hailman 1973). VARIATION WITHIN SPECIES Gaits often vary within a species (Kunkel 1962, Hailman 1973, Schwartz 1964, Gobeil 1968, Eliot in Bent 1968:669-670, this study). As an addi- tional example, I have seen Common Grackles (Quisc&s quiscula) hop in contrast to their usual walk. -

A Guide to the Birds of Barrow Island
A Guide to the Birds of Barrow Island Operated by Chevron Australia This document has been printed by a Sustainable Green Printer on stock that is certified carbon in joint venture with neutral and is Forestry Stewardship Council (FSC) mix certified, ensuring fibres are sourced from certified and well managed forests. The stock 55% recycled (30% pre consumer, 25% post- Cert no. L2/0011.2010 consumer) and has an ISO 14001 Environmental Certification. ISBN 978-0-9871120-1-9 Gorgon Project Osaka Gas | Tokyo Gas | Chubu Electric Power Chevron’s Policy on Working in Sensitive Areas Protecting the safety and health of people and the environment is a Chevron core value. About the Authors Therefore, we: • Strive to design our facilities and conduct our operations to avoid adverse impacts to human health and to operate in an environmentally sound, reliable and Dr Dorian Moro efficient manner. • Conduct our operations responsibly in all areas, including environments with sensitive Dorian Moro works for Chevron Australia as the Terrestrial Ecologist biological characteristics. in the Australasia Strategic Business Unit. His Bachelor of Science Chevron strives to avoid or reduce significant risks and impacts our projects and (Hons) studies at La Trobe University (Victoria), focused on small operations may pose to sensitive species, habitats and ecosystems. This means that we: mammal communities in coastal areas of Victoria. His PhD (University • Integrate biodiversity into our business decision-making and management through our of Western Australia) -

Captive Management for Woodhen and LHI Currawong Associated with the Lord Howe Island Rodent Eradication Project
Taronga Conservation Society Australia Captive management for Woodhen and LHI Currawong associated with the Lord Howe Island Rodent Eradication project March 2014 This report details work that occurred in the provision of captive care of Lord Howe Island Woodhen and Lord Howe Island Currawong between July 22 and October 18 2013. It details some of the preparatory work, but concentrates on recommendations for proposed work an order of magnitude larger in 2017. Approved Version: 2.0 March 2014 Page 1 of 10 Taronga Conservation Society Australia 1. Executive Summary The Lord Howe Island Board has been granted approximately $9 million to conduct an eradication program of introduced rats and mice from Lord Howe Island (LHI). This work is jointly funded through the Australian Government's Department of Environment, and the NSW Government's Environmental Trust and was announced on July 15 2012. The eradication will be via the dense distribution of a bait containing brodifacoum in a single 100 day baiting operation. Ship rats are implicated in the extinction of at least five endemic birds and at least 13 invertebrates. They are also a recognised threat to at least 13 other bird species, 2 reptiles, 51 plant species, 12 vegetation communities and numerous threatened invertebrates1. Taronga Conservation Society Australia (Taronga) was contacted in July 2009 about potential involvement in the program. It initially played an advisory role, though with the clear intention of operational involvement should the funding application be successful. A detailed risk assessment is presented which determines the risks to the environment (including wildlife, freshwater and marine habitats), humans, livestock and pets. -

Gymnorhina Tibicen Global Invasive
FULL ACCOUNT FOR: Gymnorhina tibicen Gymnorhina tibicen System: Terrestrial Kingdom Phylum Class Order Family Animalia Chordata Aves Passeriformes Cracticidae Common name Synonym Coracias tibicen Similar species Grallina cyanoleuca, Cracticus nigrogularis Summary Gymnorhina tibicen (the Australian magpie) was originally introduced to New Zealand around the 1860s in an attempt to combat pastoral pests. It is known to be extremely territorial, especially during the breeding season, and is known to assault other avian species and even humans. Magpies potentially threaten a number of indigenous avian fauna, as well as putting humans at risk of injury. view this species on IUCN Red List Species Description The Australian magpie (Gymnorhina tibicen), is a medium-sized ground-feeding passerine found throughout much of the Australian continent. They range from 36-44cm in length and weigh 280-340g with black and white plumage, black heads, wings and underparts together with white napes and shoulders (NRC, 1998). The iris of the adult magpie is red, whereas the juveniles' eyes are dark brown in colour. Minor differences exist between the male and female magpies, though in general, magpies are not considered to be sexually dimorphic (Simpson et al., 1993). Notes Although Australian magpies (Gymnorhina tibicen) have been known to have detrimental impacts on some birds, they can actually be beneficial to others. They do this by attacking and displacing common avian predators, such as harrier hawks and ravens, which in turn provides safe nesting grounds for a number of rural birds (Morgan et al, 2005). Lifecycle Stages The average life span of the Australian magpie (Gymnorhina tibicen) has not been studied in detail, but is estimated to be around 24 years, with some individuals living up to 30 years of age (Reilly, 1988). -

Grand Australia Part Ii: Queensland, Victoria & Plains-Wanderer
GRAND AUSTRALIA PART II: QUEENSLAND, VICTORIA & PLAINS-WANDERER OCTOBER 15–NOVEMBER 1, 2018 Southern Cassowary LEADER: DION HOBCROFT LIST COMPILED BY: DION HOBCROFT VICTOR EMANUEL NATURE TOURS, INC. 2525 WALLINGWOOD DRIVE, SUITE 1003 AUSTIN, TEXAS 78746 WWW.VENTBIRD.COM GRAND AUSTRALIA PART II By Dion Hobcroft Few birds are as brilliant (in an opposite complementary fashion) as a male Australian King-parrot. On Part II of our Grand Australia tour, we were joined by six new participants. We had a magnificent start finding a handsome male Koala in near record time, and he posed well for us. With friend Duncan in the “monster bus” named “Vince,” we birded through the Kerry Valley and the country towns of Beaudesert and Canungra. Visiting several sites, we soon racked up a bird list of some 90 species with highlights including two Black-necked Storks, a Swamp Harrier, a Comb-crested Jacana male attending recently fledged chicks, a single Latham’s Snipe, colorful Scaly-breasted Lorikeets and Pale-headed Rosellas, a pair of obliging Speckled Warblers, beautiful Scarlet Myzomela and much more. It had been raining heavily at O’Reilly’s for nearly a fortnight, and our arrival was exquisitely timed for a break in the gloom as blue sky started to dominate. Pretty-faced Wallaby was a good marsupial, and at lunch we were joined by a spectacular male Eastern Water Dragon. Before breakfast we wandered along the trail system adjacent to the lodge and were joined by many new birds providing unbelievable close views and photographic chances. Wonga Pigeon and Bassian Thrush were two immediate good sightings followed closely by Albert’s Lyrebird, female Paradise Riflebird, Green Catbird, Regent Bowerbird, Australian Logrunner, three species of scrubwren, and a male Rose Robin amongst others. -

The Australian Raven (Corvus Coronoides) in Metropolitan Perth
Edith Cowan University Research Online Theses : Honours Theses 1997 Some aspects of the ecology of an urban Corvid : The Australian Raven (Corvus coronoides) in metropolitan Perth P. J. Stewart Edith Cowan University Follow this and additional works at: https://ro.ecu.edu.au/theses_hons Part of the Ornithology Commons Recommended Citation Stewart, P. J. (1997). Some aspects of the ecology of an urban Corvid : The Australian Raven (Corvus coronoides) in metropolitan Perth. https://ro.ecu.edu.au/theses_hons/295 This Thesis is posted at Research Online. https://ro.ecu.edu.au/theses_hons/295 Edith Cowan University Copyright Warning You may print or download ONE copy of this document for the purpose of your own research or study. The University does not authorize you to copy, communicate or otherwise make available electronically to any other person any copyright material contained on this site. You are reminded of the following: Copyright owners are entitled to take legal action against persons who infringe their copyright. A reproduction of material that is protected by copyright may be a copyright infringement. Where the reproduction of such material is done without attribution of authorship, with false attribution of authorship or the authorship is treated in a derogatory manner, this may be a breach of the author’s moral rights contained in Part IX of the Copyright Act 1968 (Cth). Courts have the power to impose a wide range of civil and criminal sanctions for infringement of copyright, infringement of moral rights and other offences under the Copyright Act 1968 (Cth). Higher penalties may apply, and higher damages may be awarded, for offences and infringements involving the conversion of material into digital or electronic form. -

The Role of Habitat Variability and Interactions Around Nesting Cavities in Shaping Urban Bird Communities
The role of habitat variability and interactions around nesting cavities in shaping urban bird communities Andrew Munro Rogers BSc, MSc Photo: A. Rogers A thesis submitted for the degree of Doctor of Philosophy at The University of Queensland in 2018 School of Biological Sciences Andrew Rogers PhD Thesis Thesis Abstract Inter-specific interactions around resources, such as nesting sites, are an important factor by which invasive species impact native communities. As resource availability varies across different environments, competition for resources and invasive species impacts around those resources change. In urban environments, changes in habitat structure and the addition of introduced species has led to significant changes in species composition and abundance, but the extent to which such changes have altered competition over resources is not well understood. Australia’s cities are relatively recent, many of them located in coastal and biodiversity-rich areas, where conservation efforts have the opportunity to benefit many species. Australia hosts a very large diversity of cavity-nesting species, across multiple families of birds and mammals. Of particular interest are cavity-breeding species that have been significantly impacted by the loss of available nesting resources in large, old, hollow- bearing trees. Cavity-breeding species have also been impacted by the addition of cavity- breeding invasive species, increasing the competition for the remaining nesting sites. The results of this additional competition have not been quantified in most cavity breeding communities in Australia. Our understanding of the importance of inter-specific interactions in shaping the outcomes of urbanization and invasion remains very limited across Australian communities. This has led to significant gaps in the understanding of the drivers of inter- specific interactions and how such interactions shape resource use in highly modified environments. -

The Evolution of Cerebrotypes in Birds
Original Paper Brain Behav Evol 2005;65:215–230 Received: June 23, 2004 Returned for revision: July 20, 2004 DOI: 10.1159/000084313 Accepted after revision: September 14, 2004 Published online: March 8, 2005 The Evolution of Cerebrotypes in Birds Andrew N. Iwaniuk Peter L. Hurd Department of Psychology, University of Alberta, Edmonton, Canada Key Words tionships among species, but there is a tendency for spe- Birds W Wulst W Nidopallium W Brainstem W Cerebellum W cies within an order to clump together. There may also Evolution W Prey capture W Cognition be a weak relationship between cerebrotype and devel- opmental differences, but two of the main clusters con- tained species with both altricial and precocial develop- Abstract mental patterns. As a whole, the groupings do agree Multivariate analyses of brain composition in mammals, with behavioral and ecological similarities among spe- amphibians and fish have revealed the evolution of ‘cer- cies. Most notably, species that share similarities in loco- ebrotypes’ that reflect specific niches and/or clades. motor behavior, mode of prey capture or cognitive abili- Here, we present the first demonstration of similar cere- ty are clustered together. The relationship between cere- brotypes in birds. Using principal component analysis brotype and behavior/ecology in birds suggests that and hierarchical clustering methods to analyze a data set future comparative studies of brain-behavior relation- of 67 species, we demonstrate that five main cerebro- ships will benefit from adopting a multivariate ap- types can be recognized. One type is dominated by galli- proach. forms and pigeons, among other species, that all share Copyright © 2005 S. -
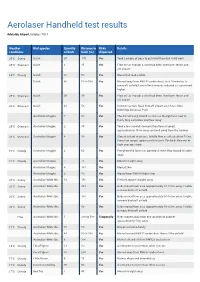
Aerolaser Handheld Test Results
Aerolaser Handheld test results Adelaide Airport October 2014 Weather Bird species Quantity Distance to Birds Details conditions of birds birds (m) dispersed 20°C Sunny Galah 20 170 Yes Took a couple of goes to get rid of them but it did work 20°C Overcast Galah 4 35 Yes Flew off as though a shot had been fired over fence and off airport 14°C Cloudy Galah 30 60 Yes Moved but took a while Galah 30 50 to 500 Yes Moved away from RWY12 undershoot, took 10 minutes to move off airfield, laser effectivenesds reduced as sun moved higher 20°C Overcast Galah 20 30 Yes Flew off as though a shot had been fired over fence and off airport 20°C Overcast Galah 40 50 Yes Instant reaction, flock flew off airport over fence intro Burbridge Business Park Australian Magpie 7 50 Yes The did not hang around as soon as the light was next to them, they carried on and flew away 20°C Overcast Australian Magpie 2 30 Yes Took a few seconds to react then flew at speed approximately 150m away and well away from the runway 20°C Overcast Australian Magpie 8 30 Yes Almost instant reactions. Initally flew as a flock about 100m. From that range I again used the laser. The birds flew out of sight over ops store 15°C Cloudy Australian Magpie 4 60 Yes Everytime the laser was pointed at them they moved straight away 15°C Cloudy Australian Magpie 7 70 Yes Moved straight away Australian Magpie 8 100 Yes Moved 24m Australian Magpie 2 50 Yes Moved from RWY05 flight strip 20°C Sunny Australian White Ibis 13 150 Yes Fled off airport straight away 20°C Sunny Australian White Ibis 250 Yes Birds moved from area approximately 50-100m away. -

Contributions to the Reproductive Effort in a Group of Plural-Breeding Pied Butcherbirds Cracticus Nigrogularis
Australian Field Ornithology 2012, 29, 169–181 Contributions to the reproductive effort in a group of plural-breeding Pied Butcherbirds Cracticus nigrogularis D.G. Gosper 39 Azure Avenue, Balnarring VIC 3926, Australia Email: [email protected] Summary. Concurrent nesting by two females from a single social group of Pied Butcherbirds Cracticus nigrogularis is described. Young fledged from two nests in the first 2 years, but breeding failed in the following two seasons after Australian Magpies C. tibicen displaced the group from its original nest-tree. The two breeding females in the Pied Butcherbird group constructed their nests and incubated synchronously in the same tree without conflict. Only the females constructed the nest and incubated the eggs. Other group members fed the females before laying and during incubation. Females begged for food using a display resembling that of juvenile Butcherbirds. Most (and probably all) members of the group fed the nestlings and fledglings, with multiple members delivering food to the young in both nests and removing or eating faecal sacs. Immatures at the beginning of their second year, with no previous experience, performed the full range of helping tasks, including the provisioning of females from the pre-laying stage. During the nestling stage, the group fed on nectar from Silky Oaks Grevillea robusta, but did not feed this to the young. Introduction The Pied Butcherbird Cracticus nigrogularis (Artamidae) may breed co-operatively (Rowley 1976; Dow 1980; Clarke 1995). Higgins et al. (2006) considered it to be an occasional co-operative breeder, although the only detailed study to date (i.e.