Grizzly Bears and Humans at Two Moth Aggregation Sites in Wyoming
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Field Notes in Support of a Revision of Hawaiian Carabidae
Field Notes in Support of a Revision of Hawaiian Carabidae James K. Liebherr* Cornell University Ithaca, NY, U.S.A. * Curator (Ret.) Cornell University Insect Collection, and Professor Emeritus 23 August 2021 Field Notes in Support of a Revision of Hawaiian Carabidae © 2021 by James K. Liebherr is licensed under Attribution-NonCommercial-NoDerivatives 4.0 International. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/4.0/ Cornell University Insect Collection John H. and Ann B. Comstock Hall Cornell University Ithaca, NY 14853-2601 U.S.A. ii Table of Contents Foreword ……………………………………………………………….…………………………. iv 00.—Introduction …………………………………………………………………..………….. 1 01.—Kauaˋi; 9-22 March, 10-16 May 1991 ……..…………………………...….... 7 02.—Hawaiˋi Island; 23 March-5 April 1991 ..…………………………..…….… 21 03.—Haleakalā; 8-13 April 1991, 29 April–9 May 1991 …………….…….… 27 04.—Oˋahu; 20–23 April, 18 May 1991 ………………………..……….…..….…..41 05.—Molokaˋi; 24-27 April 1991 …………………………………….……..….………45 06.—West Maui, 11-15 May 1992; Molokaˋi, 15-16 May 1992 ...……… 50 07.—Maui Nui, 29 April-21 May 1993 (Lānaˋi, Molokaˋi, West Maui, Haleakalā) ……………….………………………………………………………………….…….61 08.—Oˋahu, 1-15 May 1995 .………………………………….……………………..…. 89 09.—Kauaˋi, 16-22 May 1995 ………………….………………..…………..………. 104 10.—Maui + Molokaˋi 20-31 May 1997 …..……………………………………... 114 11.—Hawaiˋi Island—1–6 June 1997 …..………………………..……………….. 134 12.—Hawaiˋi Island—9–17 October 1997 …..……………………….…….….. 138 13.—Haleakalā, Maui—30 April–15 May 1998 ….…………………………... 145 14.—Haleakalā and West Maui—15–23 May 2001 …….………….………. 167 15.—Hawaiˋi Island—23–27 May 2001 …………………..…………..…………. 179 16.—Haleakalā, Maui, 14–20 May 2003 …………………………………………. 184 17.—Hawaiˋi Island, 21–25 May 2003 ………………..………………………….. 192 18.—Lānaˋi, Molokaˋi, West Maui, 10–28 May 2004 …..……..….………. -

The Integration of Campsites in Cultural Landscapes: Architectural Actions on the Catalan Coast, Spain
sustainability Article The Integration of Campsites in Cultural Landscapes: Architectural Actions on the Catalan Coast, Spain Xavier Martín * , Anna Martínez and Isabela de Rentería IAR Group, School of Architecture La Salle, Ramon Llull University, 08022 Barcelona, Spain; [email protected] (A.M.); [email protected] (I.d.R.) * Correspondence: [email protected] Received: 29 June 2020; Accepted: 6 August 2020; Published: 12 August 2020 Abstract: Over the last 60 years, the development of tourism in Spain has produced an unprecedented occupation of the territory. Urban growth, hotels and infrastructures have transformed much of the natural environment. This phenomenon has irreversibly altered conditions of regions with great landscape value, putting their cultural heritage at risk. Yet, the campsite is a model of tourist settlement based on shared living in the open natural space. It promotes minimal and temporary interventions in the territory, by means of transportable accommodations with precise occupations of place, leaving a slight footprint. Therefore, architecture contributes to affording the order and services that these individual artifacts cannot provide by themselves. In terms of slight land occupation and natural qualities preservation, the campsite has proven to be one of the most responsible tourist models. It is an opportunity for the future: a resource for landscape integration and local dynamics reactivation. We present a set of architectural actions for the integration of campsites in cultural landscapes along the Catalan coast. These are recommendations catalogued by means of a diagnosis tool that proposes strategies at different levels, from enclosure to lodgings. Focusing on end-users, this research fosters cultural identity preservation and responsible communal living in nature. -

BIOLOGICAL CONTROL of the BLACK CUTWORM, <Em>AGROTIS IPSILON</Em> (LEPIDOPTERA: NOCTUIDAE), and ENDOPHYTE MEDIATED T
University of Kentucky UKnowledge University of Kentucky Doctoral Dissertations Graduate School 2011 BIOLOGICAL CONTROL OF THE BLACK CUTWORM, AGROTIS IPSILON (LEPIDOPTERA: NOCTUIDAE), AND ENDOPHYTE MEDIATED TRITROPHIC INTERACTIONS IN TURFGRASS Andrea Jeanne Bixby-Brosi University of Kentucky, [email protected] Right click to open a feedback form in a new tab to let us know how this document benefits ou.y Recommended Citation Bixby-Brosi, Andrea Jeanne, "BIOLOGICAL CONTROL OF THE BLACK CUTWORM, AGROTIS IPSILON (LEPIDOPTERA: NOCTUIDAE), AND ENDOPHYTE MEDIATED TRITROPHIC INTERACTIONS IN TURFGRASS" (2011). University of Kentucky Doctoral Dissertations. 183. https://uknowledge.uky.edu/gradschool_diss/183 This Dissertation is brought to you for free and open access by the Graduate School at UKnowledge. It has been accepted for inclusion in University of Kentucky Doctoral Dissertations by an authorized administrator of UKnowledge. For more information, please contact [email protected]. ABSTRACT OF DISSERTATION Andrea Jeanne Bixby-Brosi The Graduate School University of Kentucky 2011 BIOLOGICAL CONTROL OF THE BLACK CUTWORM, AGROTIS IPSILON (LEPIDOPTERA: NOCTUIDAE), AND ENDOPHYTE MEDIATED TRITROPHIC INTERACTIONS IN TURFGRASS ABSTRACT OF DISSERTATION A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy in the College of Agriculture at the University of Kentucky By Andrea Jeanne Bixby-Brosi Lexington, Kentucky Director: Dr. Daniel A. Potter, Professor of Entomology Lexington, Kentucky 2011 Copyright © Andrea Jeanne Bixby-Brosi 2011 ABSTRACT OF DISSERTATION BIOLOGICAL CONTROL OF THE BLACK CUTWORM, AGROTIS IPSILON (LEPIDOPTERA: NOCTUIDAE), AND ENDOPHYTE MEDIATED TRITROPHIC INTERACTIONS IN TURFGRASS Components of successful pest management programs must be complementary and not antagonistic. This project examined interactions between natural enemies of the black cutworm, Agrotis ipsilon (Hufnagel), an important turfgrass pest, and host plant resistance by endophytic grass. -

Agrotis Ipsilon): a Glimpse on Globally Important Crop Pest
Volume 2021, issue 2, pages 36–42 30 June 2021 https://doi.org/10.33493/scivis.21.02.02 REVIEW ARTICLE An insight into black cutworm (Agrotis ipsilon): A glimpse on globally important crop pest Christopher Rodingpuia*, H. Lalthanzara Department of Life Sciences, Pachhunga University College, Mizoram University, Aizawl 796001, India Many agricultural pests create problems for farmers leading to production loss. Received 11 January 2021 Accepted 15 February 2021 Since agriculture plays an important role in the economy of a nation, controlling and management of such pests is of great importance. Cutworms are notorious *For correspondence: [email protected] agricultural pests infesting a wide variety of crops leading to production loss. Cutworms are distributed worldwide and many species of cutworms have also Contact us: been reported feeding on all kinds of crops. Agrotis ipsilon is one of the most [email protected] common cutworm species prevailing in different continents. Understanding the biology and management of these pests will be of great use for farmers. In this article, brief information on cutworms; specifically A. ipsilon species is highlighted. A brief description of host, migration, control and management of A. ipsilon are also highlighted. Keywords: Caterpillars, infestation, host, parasite, economy, control-management. Introduction Cutworms are caterpillars belonging to the order may extend up to 1 inch (2.5 cm).4 There are Lepidoptera and family Noctuidae; a large family of variations within the genera and may differ moth.1-4 Cutworms got their names from their depending on the region where they are found.1-4 feeding habit of “cutting-off”’ a seedling at ground Most common cutworm species are Agrotis level by chewing through the stem. -
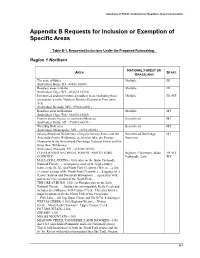
Summary of Public Comment, Appendix B
Summary of Public Comment on Roadless Area Conservation Appendix B Requests for Inclusion or Exemption of Specific Areas Table B-1. Requested Inclusions Under the Proposed Rulemaking. Region 1 Northern NATIONAL FOREST OR AREA STATE GRASSLAND The state of Idaho Multiple ID (Individual, Boise, ID - #6033.10200) Roadless areas in Idaho Multiple ID (Individual, Olga, WA - #16638.10110) Inventoried and uninventoried roadless areas (including those Multiple ID, MT encompassed in the Northern Rockies Ecosystem Protection Act) (Individual, Bemidji, MN - #7964.64351) Roadless areas in Montana Multiple MT (Individual, Olga, WA - #16638.10110) Pioneer Scenic Byway in southwest Montana Beaverhead MT (Individual, Butte, MT - #50515.64351) West Big Hole area Beaverhead MT (Individual, Minneapolis, MN - #2892.83000) Selway-Bitterroot Wilderness, along the Selway River, and the Beaverhead-Deerlodge, MT Anaconda-Pintler Wilderness, at Johnson lake, the Pioneer Bitterroot Mountains in the Beaverhead-Deerlodge National Forest and the Great Bear Wilderness (Individual, Missoula, MT - #16940.90200) CLEARWATER NATIONAL FOREST: NORTH FORK Bighorn, Clearwater, Idaho ID, MT, COUNTRY- Panhandle, Lolo WY MALLARD-LARKINS--1300 (also on the Idaho Panhandle National Forest)….encompasses most of the high country between the St. Joe and North Fork Clearwater Rivers….a low elevation section of the North Fork Clearwater….Logging sales (Lower Salmon and Dworshak Blowdown) …a potential wild and scenic river section of the North Fork... THE GREAT BURN--1301 (or Hoodoo also on the Lolo National Forest) … harbors the incomparable Kelly Creek and includes its confluence with Cayuse Creek. This area forms a major headwaters for the North Fork of the Clearwater. …Fish Lake… the Jap, Siam, Goose and Shell Creek drainages WEITAS CREEK--1306 (Bighorn-Weitas)…Weitas Creek…North Fork Clearwater. -

Sample Itineraries for Bold & Gold Expeditions All Backpacking Trips
*Note: The itineraries given therein are intended to be a sample schedule of each type of trip. These may change due to circumstances such as weather and the skill level of participants. *SAMPLE ITINERARIES FOR BOLD & GOLD EXPEDITIONS ALL BACKPACKING TRIPS DAILY TRAVEL TIME Hiking mileage will vary from trip to trip, but groups will cover between 3-10 miles a day *SAMPLE ITINERARY Day 1: Arrive at base camp, gear check, safety briefing, lunch, shuttle to trail head campsite Days 2-4: Hike to new camping destination, explore around their campsite, cook dinner, enjoy team-building activities Day 3, 4 or 5: Layover Day: a chance to camp at the same spot 2 nights in a row. Day hiking, exploring the area, outdoor skills, games FOR 1-WEEK TRIPS: Day 6-7: Final backpacking day Day 8: Return to Basecamp in Seattle, unpack gear, closing circle FOR 2-WEEK TRIPS: Days 6-7: Participants continue to learn about each other through team-building activities and take turns being “leader of the day” on the trail Day 8: A staff member will meet the group at a front country campsite with a re-ration of food and supplies Days 9-13: Participants will take on more responsibility and leadership, practicing Leave No Trace principles, cooking meals for the group and navigating the terrain. There will be another layover day Day 14: Final backpacking day Day 15: Return to Basecamp in Seattle, unpack gear, closing circle 1 WEEK TRIPS: 2 WEEK TRIPS: • Backpacking & Fishing • American Alps • Backpacking & Yoga • Backpacking & Leadership • Beyond City Limits • Backpacks, Beaches & Boulders** • Call of the North Cascades • Backpacks & Kayaks*** • Cascade Challenge • Mountain & Music • Fierce & Fabulous • Powerful Gifts**** • Make A Scene: Art & Backpacking • Olympic Coastal Backpacking • Olympic Coastal Challenge • Poets & Peaks • She-Nannigans! *Note: The itineraries given therein are intended to be a sample schedule of each type of trip. -

Region Forest Roadless Name GIS Acres 1 Beaverhead-Deerlodge
These acres were calculated from GIS data Available on the Forest Service Roadless website for the 2001 Roadless EIS. The data was downloaded on 8/24/2011 by Suzanne Johnson WO Minerals & Geology‐ GIS/Database Specialist. It was discovered that the Santa Fe NF in NM has errors. This spreadsheet holds the corrected data from the Santa Fe NF. The GIS data was downloaded from the eGIS data center SDE instance on 8/25/2011 Region Forest Roadless Name GIS Acres 1 Beaverhead‐Deerlodge Anderson Mountain 31,500.98 1 Beaverhead‐Deerlodge Basin Creek 9,499.51 1 Beaverhead‐Deerlodge Bear Creek 8,122.88 1 Beaverhead‐Deerlodge Beaver Lake 11,862.81 1 Beaverhead‐Deerlodge Big Horn Mountain 50,845.85 1 Beaverhead‐Deerlodge Black Butte 39,160.06 1 Beaverhead‐Deerlodge Call Mountain 8,795.54 1 Beaverhead‐Deerlodge Cattle Gulch 19,390.45 1 Beaverhead‐Deerlodge Cherry Lakes 19,945.49 1 Beaverhead‐Deerlodge Dixon Mountain 3,674.46 1 Beaverhead‐Deerlodge East Pioneer 145,082.05 1 Beaverhead‐Deerlodge Electric Peak 17,997.26 1 Beaverhead‐Deerlodge Emerine 14,282.26 1 Beaverhead‐Deerlodge Fleecer 31,585.50 1 Beaverhead‐Deerlodge Flint Range / Dolus Lakes 59,213.30 1 Beaverhead‐Deerlodge Four Eyes Canyon 7,029.38 1 Beaverhead‐Deerlodge Fred Burr 5,814.01 1 Beaverhead‐Deerlodge Freezeout Mountain 97,304.68 1 Beaverhead‐Deerlodge Garfield Mountain 41,891.22 1 Beaverhead‐Deerlodge Goat Mountain 9,347.87 1 Beaverhead‐Deerlodge Granulated Mountain 14,950.11 1 Beaverhead‐Deerlodge Highlands 20,043.87 1 Beaverhead‐Deerlodge Italian Peak 90,401.31 1 Beaverhead‐Deerlodge Lone Butte 13,725.16 1 Beaverhead‐Deerlodge Mckenzie Canyon 33,350.48 1 Beaverhead‐Deerlodge Middle Mtn. -

Draft Regulations
Draft 4-19-2021.2 CHAPTER 47 GRAY WOLF HUNTING SEASONS Section 1. Authority. This regulation is promulgated by the authority of Wyoming Statutes § 23-1-101, § 23-1-102, § 23-1-108, § 23-1-302, § 23-1-304, § 23-2-104, § 23-2-303, § 23-3-111, § 23-3-115 and § 23-3-304. Section 2. Definitions. In addition to the definitions set forth in Title 23 of the Wyoming Statutes and Chapter 2, General Hunting Regulation, the Commission also adopts the following definition for the purpose of this chapter: (a) “Mortality” means any legal or illegal human-caused gray wolf death that occurs during an open gray wolf hunting season, excluding gray wolves taken by the Department, gray wolves taken under the authority of W.S. § 23-3-115 (c), gray wolves taken under Lethal Take Permits and known natural and accidental gray wolf deaths. Section 3. Hunting Seasons Established. There shall be open seasons for the hunting of gray wolves as set forth in this regulation. This regulation shall go into effect from and after the date gray wolves are removed from the federal list of experimental nonessential populations, endangered species or threatened species in Wyoming. This regulation shall remain in effect until modified or repealed by the Commission. Section 4. Hunting Regulations for Gray Wolves Designated as Trophy Game Animals. (a) All gray wolves within the Wolf Trophy Game Management Area and Seasonal Wolf Trophy Game Management Area as described in W.S. §23-1-101(a) (xii) (B) (I) and (II) are designated as trophy game animals. -

Family Electric Campground
North LEGEND POTATO CREEK STATE PARK Road Hiking Trail Family Electric Campground Fire Ring Water Filling Station 29 27 118 117 30 25 120 115 *Comfort Station 31 23 113 122 111 33 28 26 22 119 116 Mountain Biking 32 24 123 121 114 110 21 34 125 124 112 108 Vault Toilet 35 20 19 36 127 126 109 107 Beach 37 18 17 39 129 128 106 105 41 38 15 Accessible 16 130 104 40 131 103 11 42 5 7 9 14 13 132 102 Playground 134 101 133 General Store 1 2 3 4 6 8 10 136 99 100 12 135 138 97 98 Camp Gatehouse 51 50 48 46 44 43 94 53 137 92 rail 5 90 96 T 54 139 Dumping Station 88 95 56 52 49 47 45 55 86 91 93 87 89 Parking 57 59 84 85 Nature Center 58 61 82 83 60 Campground Host 63 81 62 65 80 Playfield Drinking Fountain 64 79 67 Emergency Call Box 66 78 77 69 68 76 73 75 Campfire Boat Rental 70 74 71 Circle *All comfort stations are accessible. 72 To Trail 1 212 211 214 209 215 207 213 well house 210 205 217 203 216 208 202 206 269 268 218 201 271 266 204 264 141 220 200 273 T 270 267 263 ra 140 222 229 275 272 265 il 143 227 228 274 262 5 142 219 277 145 221 276 287 261 223 224 225 226 279 281 283 285 144 198 199 230 259 146 184 286 260 196 278 280 282 284 148 147 183 197 231 232 185 182 194 258 257 149 186 181 195 192 233 234 256 1 150 187 180 193 255 151 188 179 235 152 189 191 237 254 253 153 190 236 239 154 177 250 252 242 178 248 155 175 238 243 246 251 157 173 176 2 156 249 174 241 247 159 244 245 • 158 170 172 l 171 i 160 161 168 ra 166 169 T 162 167 e r 163 165 to 164 3 S l ra e Trail Beach n e G Cabins 21 20 19 18 17 16 il 4 Tra . -

Guide to the Willows of Shoshone National Forest
United States Department of Agriculture Guide to the Willows Forest Service Rocky Mountain Research Station of Shoshone National General Technical Report RMRS-GTR-83 Forest October 2001 Walter Fertig Stuart Markow Natural Resources Conservation Service Cody Conservation District Abstract Fertig, Walter; Markow, Stuart. 2001. Guide to the willows of Shoshone National Forest. Gen. Tech. Rep. RMRS-GTR-83. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 79 p. Correct identification of willow species is an important part of land management. This guide describes the 29 willows that are known to occur on the Shoshone National Forest, Wyoming. Keys to pistillate catkins and leaf morphology are included with illustrations and plant descriptions. Key words: Salix, willows, Shoshone National Forest, identification The Authors Walter Fertig has been Heritage Botanist with the University of Wyoming’s Natural Diversity Database (WYNDD) since 1992. He has conducted rare plant surveys and natural areas inventories throughout Wyoming, with an emphasis on the desert basins of southwest Wyoming and the montane and alpine regions of the Wind River and Absaroka ranges. Fertig is the author of the Wyoming Rare Plant Field Guide, and has written over 100 technical reports on rare plants of the State. Stuart Markow received his Masters Degree in botany from the University of Wyoming in 1993 for his floristic survey of the Targhee National Forest in Idaho and Wyoming. He is currently a Botanical Consultant with a research emphasis on the montane flora of the Greater Yellowstone area and the taxonomy of grasses. Acknowledgments Sincere thanks are extended to Kent Houston and Dave Henry of the Shoshone National Forest for providing Forest Service funding for this project. -

February 2, 2012 Ms. Carrie Christman Shoshone National
February 2, 2012 Ms. Carrie Christman Shoshone National Forest 808 Meadow Lane Ave. Cody, WY 82414 Dear Carrie, On behalf of the Wyoming Outdoor Council, I appreciate the opportunity to submit comments on the proposed draft forest plan. Founded in 1967, the Wyoming Outdoor Council is Wyoming’s oldest, independent statewide conservation organization. We work to safeguard public lands, wildlife and environmental quality in Wyoming. The majority of our members live in Wyoming and most of them do so in large part for the many recreational opportunities our incredible public lands offer. We have participated in the Shoshone’s plan revision process since the summer of 2005, when the plan revision first began. We’ve focused our advocacy efforts on a revised forest plan that safeguards the self-identified niche of the Shoshone as a wild, backcountry forest. The Outdoor Council is aware that the Forest Service is tasked with managing the national forests in the system for multiple uses. Within the system, however, each forest has a niche. Since the beginning of the revision process, the Shoshone has correctly identified its niche as that of a backcountry forest.1 With more than half its lands managed as wilderness, and some additional 30 percent backcountry, the Shoshone is unique. With encroaching development and a growing human population, there are fewer and fewer places in the world that are able to offer the kind of scenery, solitude and expansive backcountry experiences the Shoshone provides. It is these qualities we urge the Forest Service to celebrate and safeguard, not only for the benefit of wildlife and natural resources, but also to ensure local communities continue to benefit from the jobs and revenue created as a result of maintaining backcountry areas. -

Lander Region Angler Newsletter
Wyoming Game and Fish Department Lander Region Angler Newsletter Inside this issue: South Fork Little 1 Wind River Surveys Boysen Reservoir 3 Surveys 2017 Little and Middle 4 Popo Agie Sampling Photo Credit Bill Segraves Tiger Trout and 5 Fish Management in the Lander Region Kokanee Welcome to the 2017 Lander Region Angler Newsletter! We had another great year Wind River Sauger 6 and inside this issue you’ll find updates from our work in 2017. As always, please feel Population Rebound free to contact us with any comments or questions about the aquatic resources in the region. Your input is important to us as we manage these resources for you, the people Kids Fishing Day and 8 of Wyoming. You’ll find our contact info on the last page of this newsletter. Education Torrey, Ring, and 9 South Fork Little Wind River Surveys Trail Lakes From late-August to early-September, the Wyoming Game and Fish Department Know the 11 (WGFD) conducted fisheries surveys over a 9-day period in the South Fork Little Regulations Wind River drainage in the Wind River Mountains. The South Fork Little Wind River drainage can be accessed using the Bears Ears Trailhead out of Dickinson Park in the Important Dates in 12 Shoshone National Forest. The fisheries surveys had two primary objectives: 1) col- 2018 lect genetic samples to assist with native Yellowstone Cutthroat Trout conservation; and 2) evaluate sport fisheries, particularly those that are helicopter-stocked. Cutthroat trout genetic samples were collected from the South Fork Little Wind River and the Baptiste and Grave creek drainages to determine their source (native Yellowstone Cutthroat Trout or an introduced nonnative spe- cies) and genetic purity.