For Peer Review Only 15 Wide Formal Taxonomic Recognition (Mauyzumi and Ohba, 2004)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Haseltonia Articles and Authors.Xlsx
ABCDEFG 1 CSSA "HASELTONIA" ARTICLE TITLES #1 1993–#26 2019 AUTHOR(S) R ISSUE(S) PAGES KEY WORD 1 KEY WORD 2 2 A Cactus Database for the State of Baja California, Mexico Resendiz Ruiz, María Elena 2000 7 97-99 BajaCalifornia Database A First Record of Yucca aloifolia L. (Agavaceae/Asparagaceae) Naturalized Smith, Gideon F, Figueiredo, 3 in South Africa with Notes on its uses and Reproductive Biology Estrela & Crouch, Neil R 2012 17 87-93 Yucca Fotinos, Tonya D, Clase, Teodoro, Veloz, Alberto, Jimenez, Francisco, Griffith, M A Minimally Invasive, Automated Procedure for DNA Extraction from Patrick & Wettberg, Eric JB 4 Epidermal Peels of Succulent Cacti (Cactaceae) von 2016 22 46-47 Cacti DNA 5 A Morphological Phylogeny of the Genus Conophytum N.E.Br. (Aizoaceae) Opel, Matthew R 2005 11 53-77 Conophytum 6 A New Account of Echidnopsis Hook. F. (Asclepiadaceae: Stapeliae) Plowes, Darrel CH 1993 1 65-85 Echidnopsis 7 A New Cholla (Cactaceae) from Baja California, Mexico Rebman, Jon P 1998 6 17-21 Cylindropuntia 8 A New Combination in the genus Agave Etter, Julia & Kristen, Martin 2006 12 70 Agave A New Series of the Genus Opuntia Mill. (Opuntieae, Opuntioideae, Oakley, Luis & Kiesling, 9 Cactaceae) from Austral South America Roberto 2016 22 22-30 Opuntia McCoy, Tom & Newton, 10 A New Shrubby Species of Aloe in the Imatong Mountains, Southern Sudan Leonard E 2014 19 64-65 Aloe 11 A New Species of Aloe on the Ethiopia-Sudan Border Newton, Leonard E 2002 9 14-16 Aloe A new species of Ceropegia sect. -

Hylotelephium National Protocol: NP/HYL/1.Rev ______Botanical Taxon: Hylotelephium Telephium (L.) H
Hylotelephium National protocol: NP/HYL/1.rev ____________________________________________________________________ Botanical taxon: Hylotelephium telephium (L.) H. Ohba x H. spectabile (Boreau) H. Ohba (syn. Sedum telephium L. x S. spectabile Boreau) Common Name (when known): Sedum Date of preparation of NP: 28-08-2012 Date of revision of NP: 01-01-2020 NP revised by: W.A. Wietsma Sample to be examined: VEGETATIVE Number of growing cycles: 1 year Closing date for applications: 1/12 Submission date/period: 1/4 - 30/4 Seed/Plant Quantity: 24 young plants able to show all their characteristics during the first year of examination Special conditions sample: None Test station address: Naktuinbouw, Sotaweg 22, 2371 AA, Roelofarendsveen Name: Team Support Variety Testing Department E-mail: [email protected] List of grouping characteristics: NO, (if yes put as annex) Minimum number of plants in trial: Vegetative: 20 Seed: not appl. Minimum number of plants observed by measuring or counting: Vegetative: 1 Seed: not appl. Give description of when observations on the flower should take place: At full flowering Give description of when/where observations on the leaf should take place: At full flowering Give description of when/where the other observations should take place: At full flowering Test will take place: OUTDOOR Uniformity: A population standard of 1% with an acceptance probability of at least 95%. Number of Off-types allowed: one off-type allowed in a sample size of 24 Table of characteristics: PRESENT (see annex) (if present, please annex the table of characteristics and explanations) Literature: PRESENT (when present, please annex to this document) Page 1 of 3 Table of characteristics Hylotelephium 1. -

Sedum Society Newsletter(130) Pp
Open Research Online The Open University’s repository of research publications and other research outputs Kalanchoe arborescens - a Madagascan giant Journal Item How to cite: Walker, Colin (2019). Kalanchoe arborescens - a Madagascan giant. Sedum Society Newsletter(130) pp. 81–84. For guidance on citations see FAQs. c [not recorded] https://creativecommons.org/licenses/by-nc-nd/4.0/ Version: Version of Record Copyright and Moral Rights for the articles on this site are retained by the individual authors and/or other copyright owners. For more information on Open Research Online’s data policy on reuse of materials please consult the policies page. oro.open.ac.uk NUMBER 130 SEDUM SOCIETY NEWSLETTER JULY 2019 FRONT COVER Roy Mottram kindly supplied: “The Diet” copy of this Japanese herbal which is sharp and crisp (see page 97). “I counted the plates, and this copy is complete with 200 plates, in 8 parts, bound here in 2 vols. I checked for another Sedum but none are Established April 1987, now ending our present, so Maximowicz was basing his 32nd year. S. kagamontanum on this same plate, Subscriptions run from October to the following September. Anyone requesting translating the location as Mt. Kaga and to join after June, unless there is a special citing t.40 incorrectly. The "t.43" plate request, will receive his or her first number is also wrong. It is actually t.33 of Newsletter in October. If you do not the whole work, or Vol.2 t.8. The book is receive your copy by the 10th of April, July or October, or the 15th January, then bound back to front [by Western standards] please write to the editor: Ray as in all Japanese books of the day.” RM. -

A Numerical Taxonomy of the Genus Sedum L., from Pakistan and Kashmir
Pak. J. Bot., 43(2): 753-758, 2011. A NUMERICAL TAXONOMY OF THE GENUS SEDUM L., FROM PAKISTAN AND KASHMIR GHULAM RASOOL SARWAR AND MUHAMMAD QAISER1 Centre for Plant Conservation, University of Karachi, Karachi-75270, Pakistan 1Federal Urdu University of Arts, Science and Technology, Gulshan-e-Iqbal, Karachi, Pakistan Corresponding author e-mail: [email protected] Abstract The phenetic relationship between the species of the genus Sedum L. was investigated. Data from macro and micromorphology, including pollen and seed morphology, chemistry and distribution pattern was utilized. Two distinct groups of taxa are recognized from which one group comprises S. multicaule Wall. ex Lindl., and S. hispanicum L., while other group consists of S. trullipetalum Hook. f. & Thomson S. fischeri Raym. - Hamet and S. oreades (Decne.) Raym. – Hamet. A key of the taxa is provided and distribution maps of the species are also presented. Introduction The importance of statistical methods in biological sciences can hardly be over emphasized. Many workers were of the view that numerical analysis is the comprehensive way of evaluating and analyzing the data and producing a phenetic classification (Sokal & Sneath, 1963; Mc Neil et al., 1969; Sneath & Sokal, 1973). Cronquist (1964) debated the same issue and emphasized that data collected subjectively should not become objective, when analyzed and presented objectively. Considerable work has been done on family Crassulaceae to elucidate the phylogenetic relationship. Uhl (1963) studied the phylogeny of the family Crassulaceae. Hart (1982) analyzed the relationships within the subfamily Sedoideae. Mes (1995) did the phylogenetic analysis of 22 taxa belonging to Sempervivoideae and five taxa of Sedoideae. -

NJ Native Plants - USDA
NJ Native Plants - USDA Scientific Name Common Name N/I Family Category National Wetland Indicator Status Thermopsis villosa Aaron's rod N Fabaceae Dicot Rubus depavitus Aberdeen dewberry N Rosaceae Dicot Artemisia absinthium absinthium I Asteraceae Dicot Aplectrum hyemale Adam and Eve N Orchidaceae Monocot FAC-, FACW Yucca filamentosa Adam's needle N Agavaceae Monocot Gentianella quinquefolia agueweed N Gentianaceae Dicot FAC, FACW- Rhamnus alnifolia alderleaf buckthorn N Rhamnaceae Dicot FACU, OBL Medicago sativa alfalfa I Fabaceae Dicot Ranunculus cymbalaria alkali buttercup N Ranunculaceae Dicot OBL Rubus allegheniensis Allegheny blackberry N Rosaceae Dicot UPL, FACW Hieracium paniculatum Allegheny hawkweed N Asteraceae Dicot Mimulus ringens Allegheny monkeyflower N Scrophulariaceae Dicot OBL Ranunculus allegheniensis Allegheny Mountain buttercup N Ranunculaceae Dicot FACU, FAC Prunus alleghaniensis Allegheny plum N Rosaceae Dicot UPL, NI Amelanchier laevis Allegheny serviceberry N Rosaceae Dicot Hylotelephium telephioides Allegheny stonecrop N Crassulaceae Dicot Adlumia fungosa allegheny vine N Fumariaceae Dicot Centaurea transalpina alpine knapweed N Asteraceae Dicot Potamogeton alpinus alpine pondweed N Potamogetonaceae Monocot OBL Viola labradorica alpine violet N Violaceae Dicot FAC Trifolium hybridum alsike clover I Fabaceae Dicot FACU-, FAC Cornus alternifolia alternateleaf dogwood N Cornaceae Dicot Strophostyles helvola amberique-bean N Fabaceae Dicot Puccinellia americana American alkaligrass N Poaceae Monocot Heuchera americana -
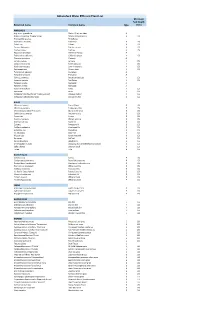
Abbotsford Water Efficient Plant List Minimum Soil Depth Botanical Name Common Name Type (Mm)
Abbotsford Water Efficient Plant List Minimum Soil Depth Botanical name Common name type (mm) ANNUALS Argemone grandifolia Statice/Sea Lavendar A Begonia x hybrida 'Dragon Wings' Dragon Wing begonia A 150 Bracteantha species Strawflower Calendula officinalis Calendula A 150 Coleus ssp. Coleus A 150 Cosmos bipinnatus Garden cosmos A 150 Cuphea llavea Cuphea A 150 Dyssocua tenuiloba Dahlberg Daisey A Eschscholzia californica California poppy A 150 Gazania spendens Gazania A Lantana camara Lantana A 150 Lobularia maritima Sweet alyssum A 150 Nigella damascena Love-in-the-mist A 150 Oesteopermum African daisy A 150 Pelargonium species Geranium A Portulaca oleracea Portulaca A Salvia guaranitica Anise-scented sage A 150 Scaevola aemula Fan flower A 150 Targetes erecta Marogold A Targetes erecta Marogold A Viola x wittrockiana Pansy A 150 Zinnia ssp. Zinnia A 150 Verbascum bombyciferum 'Arctic Summer' Broussa mullein A 150 Verbascum phlomoides 'Spica' Orange mullein A 150 BULBS Allium christophii Star of Persia B 150 Allium karataviense Turkestan onion B 150 Chionodoxa forbesii 'Pink Giant' Glory of the snow B 150 Colchicum autumnale Autumn crocus B 150 Crocus ssp. Crocus B 150 Eranthis hyemalis Winter aconite B 150 Erythronium ssp. Fawn lily B 150 Eucomis Pineapple lily B 150 Fritillaria meleagris Checkered lily B 150 Galanthus ssp. Snowdrop B 150 Iris reticulata Dwarf iris B 150 Muscari ssp. Grape hyacinth B 150 Narcissus Daffodil B 150 Nerine bowdenii Bowden lily B 150 Ornithogalum nutans Drooping Star of Bethlehem/Silverbells B 150 Scilla -

Universidad Nacional Autónoma De México Facultad De Estudios
Universidad Nacional Autónoma de México Facultad de Estudios Superiores Zaragoza “LAS CRASULÁCEAS DEL VALLE DEL MEZQUITAL’’ Tesis de licenciatura que para obtener el título de Biólogo presentan: Gabriela de Jesús Espino Ortega Luis Emilio de la Cruz López Área de Botánica Director de tesis: M. en C. Balbina Vázquez Benítez. _____________________________ Marzo 2009 UNAM – Dirección General de Bibliotecas Tesis Digitales Restricciones de uso DERECHOS RESERVADOS © PROHIBIDA SU REPRODUCCIÓN TOTAL O PARCIAL Todo el material contenido en esta tesis esta protegido por la Ley Federal del Derecho de Autor (LFDA) de los Estados Unidos Mexicanos (México). El uso de imágenes, fragmentos de videos, y demás material que sea objeto de protección de los derechos de autor, será exclusivamente para fines educativos e informativos y deberá citar la fuente donde la obtuvo mencionando el autor o autores. Cualquier uso distinto como el lucro, reproducción, edición o modificación, será perseguido y sancionado por el respectivo titular de los Derechos de Autor. Agradecimientos A la UNAM por brindarme la oportunidad de desarrollarme mejor tanto en el sentido humano como en el profesional. Por hacer la diferencia en mi vida. A la FES Zaragoza y a las personas que en ella laboran, por ser mi casa de estudios. Al grupo de sinodales formado por M. en C. Ramiro Ríos Gómez, M. en C. Balbina Vázquez Benítez, M. en C. Efraín Ángeles Cervantes, M. en C. Carlos Castillejos Cruz y Florencia Becerril Cruz, por las observaciones hechas pertinentemente para la mejora de este trabajo, por los comentarios y el tiempo dedicado al mismo. Por su amistad A la Maestra Balbina con quien estaré siempre agradecida porque trabajó con mucho entusiasmo en nuestro trabajo. -

The Rare Plant Register of South Northumberland (VC67) 2010 Quentin J
The Rare Plant Register of South Northumberland (VC67) 2010 Quentin J. Groom and A. John Richards Introduction The Vice-County Rare Plant Registers are an initiative of the Botanical Society of the British Isles to summarise the status of rare and conservation-worthy plants in each vice-county. The intention is to create an up-to-date summary of the sites of rare plants and their status at these sites. Rare Plant Registers intend to identify gaps in our knowledge, aid conservation efforts and encourage monitoring of our rare plants. Criteria for Inclusion The guidelines of the BSBI were followed in the production of this Rare Plant Register. All native vascular plants with a national status of “rare” (found in 1-15 hectads in Britain) or “scarce” (found in 16-100 hectads in Britain) are included even if that species is not native to South Northumberland. In addition, all native species locally rare or scarce in South Northumberland are included, as are extinct native species. These guidelines were occasionally relaxed to include some local specialities and hybrids of note. We would have liked to restrict the list to current sites for each species. However, in many cases, there is too little up-to-date information to make this possible. The listed sites are those where the species might still exist or has existed recently. In most cases, a site is included if a species has been recorded there since 1970. Sites without detailed locality information or of dubious provenance are not included. Where possible, we have tried to show the known history of a site by noting the date of first and last record. -

PLANTS of PEEBLESSHIRE (Vice-County 78)
PLANTS OF PEEBLESSHIRE (Vice-county 78) A CHECKLIST OF FLOWERING PLANTS AND FERNS David J McCosh 2012 Cover photograph: Sedum villosum, FJ Roberts Cover design: L Cranmer Copyright DJ McCosh Privately published DJ McCosh Holt Norfolk 2012 2 Neidpath Castle Its rocks and grassland are home to scarce plants 3 4 Contents Introduction 1 History of Plant Recording 1 Geographical Scope and Physical Features 2 Characteristics of the Flora 3 Sources referred to 5 Conventions, Initials and Abbreviations 6 Plant List 9 Index of Genera 101 5 Peeblesshire (v-c 78), showing main geographical features 6 Introduction This book summarises current knowledge about the distribution of wild flowers in Peeblesshire. It is largely the fruit of many pleasant hours of botanising by the author and a few others and as such reflects their particular interests. History of Plant Recording Peeblesshire is thinly populated and has had few resident botanists to record its flora. Also its upland terrain held little in the way of dramatic features or geology to attract outside botanists. Consequently the first list of the county’s flora with any pretension to completeness only became available in 1925 with the publication of the History of Peeblesshire (Eds, JW Buchan and H Paton). For this FRS Balfour and AB Jackson provided a chapter on the county’s flora which included a list of all the species known to occur. The first records were made by Dr A Pennecuik in 1715. He gave localities for 30 species and listed 8 others, most of which are still to be found. Thereafter for some 140 years the only evidence of interest is a few specimens in the national herbaria and scattered records in Lightfoot (1778), Watson (1837) and The New Statistical Account (1834-45). -

Threats to Australia's Grazing Industries by Garden
final report Project Code: NBP.357 Prepared by: Jenny Barker, Rod Randall,Tony Grice Co-operative Research Centre for Australian Weed Management Date published: May 2006 ISBN: 1 74036 781 2 PUBLISHED BY Meat and Livestock Australia Limited Locked Bag 991 NORTH SYDNEY NSW 2059 Weeds of the future? Threats to Australia’s grazing industries by garden plants Meat & Livestock Australia acknowledges the matching funds provided by the Australian Government to support the research and development detailed in this publication. This publication is published by Meat & Livestock Australia Limited ABN 39 081 678 364 (MLA). Care is taken to ensure the accuracy of the information contained in this publication. However MLA cannot accept responsibility for the accuracy or completeness of the information or opinions contained in the publication. You should make your own enquiries before making decisions concerning your interests. Reproduction in whole or in part of this publication is prohibited without prior written consent of MLA. Weeds of the future? Threats to Australia’s grazing industries by garden plants Abstract This report identifies 281 introduced garden plants and 800 lower priority species that present a significant risk to Australia’s grazing industries should they naturalise. Of the 281 species: • Nearly all have been recorded overseas as agricultural or environmental weeds (or both); • More than one tenth (11%) have been recorded as noxious weeds overseas; • At least one third (33%) are toxic and may harm or even kill livestock; • Almost all have been commercially available in Australia in the last 20 years; • Over two thirds (70%) were still available from Australian nurseries in 2004; • Over two thirds (72%) are not currently recognised as weeds under either State or Commonwealth legislation. -

Invasive Alien Species in Switzerland
> Environmental studies > Organisms 29 > Invasive alien species 06 in Switzerland An inventory of alien species and their threat to biodiversity and economy in Switzerland > Environmental studies > Organisms > Invasive alien species in Switzerland An inventory of alien species and their threat to biodiversity and economy in Switzerland Mit deutscher Zusammenfassung – Avec résumé en français Published by the Federal Office for the Environment FOEN Bern, 2006 Impressum Editor Federal Office for the Environment (FOEN) FOEN is an office of the Federal Department of Environment, Transport, Energy and Communications (DETEC). Authors Rüdiger Wittenberg, CABI Bioscience Switzerland Centre, CH–2800 Delémont Marc Kenis, CABI Bioscience Switzerland Centre, CH–2800 Delémont Theo Blick, D–95503 Hummeltal Ambros Hänggi, Naturhistorisches Museum, CH–4001 Basel André Gassmann, CABI Bioscience Switzerland Centre, CH–2800 Delémont Ewald Weber, Geobotanical Institute, Swiss Federal Institute of Technology, CH–8044 Zürich FOEN consultant Hans Hosbach, Head of Section, Section Biotechnology Suggested form of citation Wittenberg, R. (ed.) (2005) An inventory of alien species and their threat to biodiversity and economy in Switzerland. CABI Bioscience Switzerland Centre report to the Swiss Agency for Environment, Forests and Landscape. The environment in practice no. 0629. Federal Office for the Environment, Bern. 155 pp. Design Ursula Nöthiger-Koch, 4813 Uerkheim Fact sheets The fact sheets are available at www.environment-switzerland.ch/uw-0629-e Pictures Cover picture: Harmonia axyridis Photo Marc Kenis, CABI Bioscience, Delémont. Orders FOEN Documentation CH-3003 Bern Fax +41 (0)31 324 02 16 [email protected] www.environment-switzerland.ch/uw-0629-e Order number and price: UW-0629-E / CHF 20.– (incl. -

The Pennsylvania State University
The Pennsylvania State University The Graduate School Department of Horticulture UTILIZING SEDUM SEEDS AS AN INSTALLATION METHOD FOR GREEN ROOFS THROUGH SEED ENHANCEMENT TECHNIQUES A Dissertation in Horticulture by Kathryn Lyn McDavid 2012 Kathryn Lyn McDavid Submitted in Partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy May 2012 The dissertation of Kathryn L. McDavid was reviewed and approved* by the following: Robert Berghage Associate Professor of Horticulture Dissertation Advisor Chair of Committee Ricky M. Bates Associate Professor of Ornamental Horticulture E. Jay Holcomb Emeritus Professor of Floriculture Robert D. Shannon Associate Professor of Agricultural Engineering Richard Marini Head and Professor of the Department of Horticulture *Signatures are on file in the Graduate School iii ABSTRACT There are many benefits to installing green roofs including reducing storm water runoff and the associated problems with overloaded sewage systems, especially in high density urban areas. Green roofs also improve rain water and air quality and reduce the effect from urban heat islands by acting as thermal damper/evaporative coolers, which decrease heating and cooling costs and the subsequent pollution. Additionally, green roofs can extend the life of a roof and create natural habitats for birds and insects. They are also aesthetically pleasing which can improve people’s psychological state and productivity levels. One of the primary limitations to increased green roof installations in the United States is the cost, specifically, the high costs of labor to install the plants on the roof. The majority of green roofs today are installed using cuttings and/or plugs, nursery containers, vegetative mats, or modules.