A Distinctive Late Triassic Microvertebrate Fissure Fauna and A
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Macroevolutionary Patterns in Rhynchocephalia: Is the Tuatara (Sphenodon Punctatus) a Living Fossil? Palaeontology, 60(3), 319-328
Herrera Flores, J., Stubbs, T., & Benton, M. (2017). Macroevolutionary patterns in Rhynchocephalia: is the tuatara (Sphenodon punctatus) a living fossil? Palaeontology, 60(3), 319-328. DOI: 10.1111/pala.12284 Publisher's PDF, also known as Version of record License (if available): CC BY Link to published version (if available): 10.1111/pala.12284 Link to publication record in Explore Bristol Research PDF-document University of Bristol - Explore Bristol Research General rights This document is made available in accordance with publisher policies. Please cite only the published version using the reference above. Full terms of use are available: http://www.bristol.ac.uk/pure/about/ebr-terms.html [Palaeontology, 2017, pp. 1–10] MACROEVOLUTIONARY PATTERNS IN RHYNCHOCEPHALIA: IS THE TUATARA (SPHENODON PUNCTATUS) A LIVING FOSSIL? by JORGEA.HERRERA-FLORES , THOMAS L. STUBBS and MICHAEL J. BENTON School of Earth Sciences, University of Bristol, Wills Memorial Building, Queens Road, Bristol, BS8 1RJ, UK; jorge.herrerafl[email protected], [email protected], [email protected] Typescript received 4 July 2016; accepted in revised form 19 January 2017 Abstract: The tuatara, Sphenodon punctatus, known from since the Triassic, rhynchocephalians had heterogeneous rates 32 small islands around New Zealand, has often been noted of morphological evolution and occupied wide mor- as a classic ‘living fossil’ because of its apparently close phospaces during the Triassic and Jurassic, and these then resemblance to its Mesozoic forebears and because of a long, declined in the Cretaceous. In particular, we demonstrate low-diversity history. This designation has been disputed that the extant tuatara underwent unusually slow lineage because of the wide diversity of Mesozoic forms and because evolution, and is morphologically conservative, being located of derived adaptations in living Sphenodon. -

Studies on Continental Late Triassic Tetrapod Biochronology. I. the Type Locality of Saturnalia Tupiniquim and the Faunal Succession in South Brazil
Journal of South American Earth Sciences 19 (2005) 205–218 www.elsevier.com/locate/jsames Studies on continental Late Triassic tetrapod biochronology. I. The type locality of Saturnalia tupiniquim and the faunal succession in south Brazil Max Cardoso Langer* Departamento de Biologia, FFCLRP, Universidade de Sa˜o Paulo (USP), Av. Bandeirantes 3900, 14040-901 Ribeira˜o Preto, SP, Brazil Received 1 November 2003; accepted 1 January 2005 Abstract Late Triassic deposits of the Parana´ Basin, Rio Grande do Sul, Brazil, encompass a single third-order, tetrapod-bearing sedimentary sequence that includes parts of the Alemoa Member (Santa Maria Formation) and the Caturrita Formation. A rich, diverse succession of terrestrial tetrapod communities is recorded in these sediments, which can be divided into at least three faunal associations. The stem- sauropodomorph Saturnalia tupiniquim was collected in the locality known as ‘Waldsanga’ near the city of Santa Maria. In that area, the deposits of the Alemoa Member yield the ‘Alemoa local fauna,’ which typifies the first association; includes the rhynchosaur Hyperodapedon, aetosaurs, and basal dinosaurs; and is coeval with the lower fauna of the Ischigualasto Formation, Bermejo Basin, NW Argentina. The second association is recorded in deposits of both the Alemoa Member and the Caturrita Formation, characterized by the rhynchosaur ‘Scaphonyx’ sulcognathus and the cynodont Exaeretodon, and correlated with the upper fauna of the Ischigualasto Formation. Various isolated outcrops of the Caturrita Formation yield tetrapod fossils that correspond to post-Ischigualastian faunas but might not belong to a single faunal association. The record of the dicynodont Jachaleria suggests correlations with the lower part of the Los Colorados Formation, NW Argentina, whereas remains of derived tritheledontid cynodonts indicate younger ages. -

Constraints on the Timescale of Animal Evolutionary History
Palaeontologia Electronica palaeo-electronica.org Constraints on the timescale of animal evolutionary history Michael J. Benton, Philip C.J. Donoghue, Robert J. Asher, Matt Friedman, Thomas J. Near, and Jakob Vinther ABSTRACT Dating the tree of life is a core endeavor in evolutionary biology. Rates of evolution are fundamental to nearly every evolutionary model and process. Rates need dates. There is much debate on the most appropriate and reasonable ways in which to date the tree of life, and recent work has highlighted some confusions and complexities that can be avoided. Whether phylogenetic trees are dated after they have been estab- lished, or as part of the process of tree finding, practitioners need to know which cali- brations to use. We emphasize the importance of identifying crown (not stem) fossils, levels of confidence in their attribution to the crown, current chronostratigraphic preci- sion, the primacy of the host geological formation and asymmetric confidence intervals. Here we present calibrations for 88 key nodes across the phylogeny of animals, rang- ing from the root of Metazoa to the last common ancestor of Homo sapiens. Close attention to detail is constantly required: for example, the classic bird-mammal date (base of crown Amniota) has often been given as 310-315 Ma; the 2014 international time scale indicates a minimum age of 318 Ma. Michael J. Benton. School of Earth Sciences, University of Bristol, Bristol, BS8 1RJ, U.K. [email protected] Philip C.J. Donoghue. School of Earth Sciences, University of Bristol, Bristol, BS8 1RJ, U.K. [email protected] Robert J. -

Dinosaur Remains from the 'Botucaraí Hill' (Caturrita Formation), Late
This article was downloaded by: [Jonathas Souza Bittencourt] On: 17 August 2012, At: 06:23 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Historical Biology: An International Journal of Paleobiology Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/ghbi20 Dinosaur remains from the ‘Botucaraí Hill’ (Caturrita Formation), Late Triassic of south Brazil, and their stratigraphic context Jonathas Souza Bittencourt a b , Átila Augusto Stock da Rosa c , Cesar Leandro Schultz d & Max Cardoso Langer a a Departamento de Biologia, Universidade de São Paulo, Av. Bandeirantes 3900, 1404-901, Ribeirão Preto (SP), Brazil b Departamento de Geologia, Universidade Federal de Minas Gerais, Av. Antônio Carlos 6670, 31270-901, Belo Horizonte (MG), Brazil c Departamento de Geociências, Universidade Federal de Santa Maria, Av. Roraima 1000, 97105-900, Santa Maria (RS), Brazil d Instituto de Geociências, Universidade Federal do Rio Grande do Sul, Av. Bento Gonçalves 9500, 91501-970, Porto Alegre (RS), Brazil Version of record first published: 17 Aug 2012 To cite this article: Jonathas Souza Bittencourt, Átila Augusto Stock da Rosa, Cesar Leandro Schultz & Max Cardoso Langer (2012): Dinosaur remains from the ‘Botucaraí Hill’ (Caturrita Formation), Late Triassic of south Brazil, and their stratigraphic context, Historical Biology: An International Journal of Paleobiology, DOI:10.1080/08912963.2012.694881 To link to this article: http://dx.doi.org/10.1080/08912963.2012.694881 PLEASE SCROLL DOWN FOR ARTICLE Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions This article may be used for research, teaching, and private study purposes. -

Mesozoic Marine Reptile Palaeobiogeography in Response to Drifting Plates
ÔØ ÅÒÙ×Ö ÔØ Mesozoic marine reptile palaeobiogeography in response to drifting plates N. Bardet, J. Falconnet, V. Fischer, A. Houssaye, S. Jouve, X. Pereda Suberbiola, A. P´erez-Garc´ıa, J.-C. Rage, P. Vincent PII: S1342-937X(14)00183-X DOI: doi: 10.1016/j.gr.2014.05.005 Reference: GR 1267 To appear in: Gondwana Research Received date: 19 November 2013 Revised date: 6 May 2014 Accepted date: 14 May 2014 Please cite this article as: Bardet, N., Falconnet, J., Fischer, V., Houssaye, A., Jouve, S., Pereda Suberbiola, X., P´erez-Garc´ıa, A., Rage, J.-C., Vincent, P., Mesozoic marine reptile palaeobiogeography in response to drifting plates, Gondwana Research (2014), doi: 10.1016/j.gr.2014.05.005 This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain. ACCEPTED MANUSCRIPT Mesozoic marine reptile palaeobiogeography in response to drifting plates To Alfred Wegener (1880-1930) Bardet N.a*, Falconnet J. a, Fischer V.b, Houssaye A.c, Jouve S.d, Pereda Suberbiola X.e, Pérez-García A.f, Rage J.-C.a and Vincent P.a,g a Sorbonne Universités CR2P, CNRS-MNHN-UPMC, Département Histoire de la Terre, Muséum National d’Histoire Naturelle, CP 38, 57 rue Cuvier, -

A Small Lepidosauromorph Reptile from the Early Triassic of Poland
A SMALL LEPIDOSAUROMORPH REPTILE FROM THE EARLY TRIASSIC OF POLAND SUSAN E. EVANS and MAGDALENA BORSUK−BIAŁYNICKA Evans, S.E. and Borsuk−Białynicka, M. 2009. A small lepidosauromorph reptile from the Early Triassic of Poland. Palaeontologia Polonica 65, 179–202. The Early Triassic karst deposits of Czatkowice quarry near Kraków, southern Poland, has yielded a diversity of fish, amphibians and small reptiles. Two of these reptiles are lepido− sauromorphs, a group otherwise very poorly represented in the Triassic record. The smaller of them, Sophineta cracoviensis gen. et sp. n., is described here. In Sophineta the unspecial− ised vertebral column is associated with the fairly derived skull structure, including the tall facial process of the maxilla, reduced lacrimal, and pleurodonty, that all resemble those of early crown−group lepidosaurs rather then stem−taxa. Cladistic analysis places this new ge− nus as the sister group of Lepidosauria, displacing the relictual Middle Jurassic genus Marmoretta and bringing the origins of Lepidosauria closer to a realistic time frame. Key words: Reptilia, Lepidosauria, Triassic, phylogeny, Czatkowice, Poland. Susan E. Evans [[email protected]], Department of Cell and Developmental Biology, Uni− versity College London, Gower Street, London, WC1E 6BT, UK. Magdalena Borsuk−Białynicka [[email protected]], Institut Paleobiologii PAN, Twarda 51/55, PL−00−818 Warszawa, Poland. Received 8 March 2006, accepted 9 January 2007 180 SUSAN E. EVANS and MAGDALENA BORSUK−BIAŁYNICKA INTRODUCTION Amongst living reptiles, lepidosaurs (snakes, lizards, amphisbaenians, and tuatara) form the largest and most successful group with more than 7 000 widely distributed species. The two main lepidosaurian clades are Rhynchocephalia (the living Sphenodon and its extinct relatives) and Squamata (lizards, snakes and amphisbaenians). -

Gondwana Vertebrate Faunas of India: Their Diversity and Intercontinental Relationships
438 Article 438 by Saswati Bandyopadhyay1* and Sanghamitra Ray2 Gondwana Vertebrate Faunas of India: Their Diversity and Intercontinental Relationships 1Geological Studies Unit, Indian Statistical Institute, 203 B. T. Road, Kolkata 700108, India; email: [email protected] 2Department of Geology and Geophysics, Indian Institute of Technology, Kharagpur 721302, India; email: [email protected] *Corresponding author (Received : 23/12/2018; Revised accepted : 11/09/2019) https://doi.org/10.18814/epiiugs/2020/020028 The twelve Gondwanan stratigraphic horizons of many extant lineages, producing highly diverse terrestrial vertebrates India have yielded varied vertebrate fossils. The oldest in the vacant niches created throughout the world due to the end- Permian extinction event. Diapsids diversified rapidly by the Middle fossil record is the Endothiodon-dominated multitaxic Triassic in to many communities of continental tetrapods, whereas Kundaram fauna, which correlates the Kundaram the non-mammalian synapsids became a minor components for the Formation with several other coeval Late Permian remainder of the Mesozoic Era. The Gondwana basins of peninsular horizons of South Africa, Zambia, Tanzania, India (Fig. 1A) aptly exemplify the diverse vertebrate faunas found Mozambique, Malawi, Madagascar and Brazil. The from the Late Palaeozoic and Mesozoic. During the last few decades much emphasis was given on explorations and excavations of Permian-Triassic transition in India is marked by vertebrate fossils in these basins which have yielded many new fossil distinct taxonomic shift and faunal characteristics and vertebrates, significant both in numbers and diversity of genera, and represented by small-sized holdover fauna of the providing information on their taphonomy, taxonomy, phylogeny, Early Triassic Panchet and Kamthi fauna. -

Final Copy 2019 10 01 Herrera
This electronic thesis or dissertation has been downloaded from Explore Bristol Research, http://research-information.bristol.ac.uk Author: Herrera Flores, Jorge Alfredo A Title: The macroevolution and macroecology of Mesozoic lepidosaurs General rights Access to the thesis is subject to the Creative Commons Attribution - NonCommercial-No Derivatives 4.0 International Public License. A copy of this may be found at https://creativecommons.org/licenses/by-nc-nd/4.0/legalcode This license sets out your rights and the restrictions that apply to your access to the thesis so it is important you read this before proceeding. Take down policy Some pages of this thesis may have been removed for copyright restrictions prior to having it been deposited in Explore Bristol Research. However, if you have discovered material within the thesis that you consider to be unlawful e.g. breaches of copyright (either yours or that of a third party) or any other law, including but not limited to those relating to patent, trademark, confidentiality, data protection, obscenity, defamation, libel, then please contact [email protected] and include the following information in your message: •Your contact details •Bibliographic details for the item, including a URL •An outline nature of the complaint Your claim will be investigated and, where appropriate, the item in question will be removed from public view as soon as possible. This electronic thesis or dissertation has been downloaded from Explore Bristol Research, http://research-information.bristol.ac.uk Author: Herrera Flores, Jorge Alfredo A Title: The macroevolution and macroecology of Mesozoic lepidosaurs General rights Access to the thesis is subject to the Creative Commons Attribution - NonCommercial-No Derivatives 4.0 International Public License. -
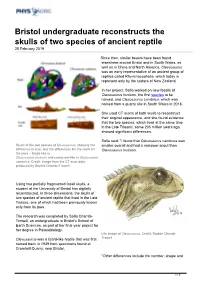
Bristol Undergraduate Reconstructs the Skulls of Two Species of Ancient Reptile 25 February 2019
Bristol undergraduate reconstructs the skulls of two species of ancient reptile 25 February 2019 Since then, similar beasts have been found elsewhere around Bristol and in South Wales, as well as in China and North America. Clevosaurus was an early representative of an ancient group of reptiles called Rhynchocephalia, which today is represent only by the tuatara of New Zealand. In her project, Sofia worked on new fossils of Clevosaurus hudsoni, the first species to be named, and Clevosaurus cambrica, which was named from a quarry site in South Wales in 2018. She used CT scans of both skulls to reconstruct their original appearance, and she found evidence that the two species, which lived at the same time in the Late Triassic, some 205 million years ago, showed significant differences. Sofia said: "I found that Clevosaurus cambrica was Skulls of the two species of Clevosaurus, showing the smaller overall and had a narrower snout than difference in size, and the differences kin the teeth kin Clevosaurus hudsoni. the jaws -- blade-like in Clevosaurus hudsoni and corkscrew-like in Clevosaurus cambrica. Credit: Image from the CT scan data, produced by Sophie Chambi-Trowell. Using two partially fragmented fossil skulls, a student at the University of Bristol has digitally reconstructed, in three-dimensions, the skulls of two species of ancient reptile that lived in the Late Triassic, one of which had been previously known only from its jaws. The research was completed by Sofia Chambi- Trowell, an undergraduate in Bristol's School of Earth Sciences, as part of her final-year project for her degree in Palaeobiology. -

The Tiny Cretaceous Stem-Bird Oculudentavis Revealed As a Bizarre Lizard
bioRxiv preprint doi: https://doi.org/10.1101/2020.08.09.243048; this version posted August 10, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The tiny Cretaceous stem-bird Oculudentavis revealed as a bizarre lizard Arnau Bolet1,2, Edward L. Stanley3, Juan D. Daza4*, J. Salvador Arias5, Andrej Čerňanský6, Marta Vidal-García7, Aaron M. Bauer8, Joseph J. Bevitt9, Adolf Peretti10, and Susan E. Evans11 Affiliations: 1 Institut Català de Paleontologia, Universitat Autònoma de Barcelona. Barcelona, Spain. 2 School of Earth Sciences, University of Bristol, Bristol, United Kingdom. 3 Department of Herpetology, Florida Museum of Natural History, Gainesville, Florida, United States. 4 Department of Biological Sciences, Sam Houston State University, Huntsville, Texas, United States. 5 Fundación Miguel Lillo, CONICET, San Miguel de Tucumán, Argentina. 6 Department of Ecology, Laboratory of Evolutionary Biology, Faculty of Natural Sciences, Comenius University in Bratislava, Bratislava, Slovakia. 7Department of Cell Biology & Anatomy, University of Calgary, Calgary, Canada. 8Department of Biology and Center for Biodiversity and Ecosystem Stewardship, Villanova University, Villanova, Pennsylvania, United States. 9Australian Centre for Neutron Scattering, Australian Nuclear Science and Technology Organisation, Sydney, Australia. 10GRS Gemresearch Swisslab AG and Peretti Museum Foundation, Meggen, Switzerland. 11Department of Cell and Developmental Biology, University College London, London, United Kingdom. *For correspondence: [email protected] 1 bioRxiv preprint doi: https://doi.org/10.1101/2020.08.09.243048; this version posted August 10, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. -

Biomechanical Assessment of Evolutionary Changes in the Lepidosaurian Skull
Biomechanical assessment of evolutionary changes in the lepidosaurian skull Mehran Moazena,1, Neil Curtisa, Paul O’Higginsb, Susan E. Evansc, and Michael J. Fagana aDepartment of Engineering, University of Hull, Hull HU6 7RX, United Kingdom; bThe Hull York Medical School, University of York, York YO10 5DD, United Kingdom; and cResearch Department of Cell and Developmental Biology, University College London, Gower Street, London WC1E 6BT, United Kingdom Edited by R. McNeill Alexander, University of Leeds, Leeds, United Kingdom, and accepted by the Editorial Board March 24, 2009 (received for review December 23, 2008) The lepidosaurian skull has long been of interest to functional mor- a synovial joint with the pterygoid, the base resting in a pit (fossa phologists and evolutionary biologists. Patterns of bone loss and columellae) on the dorsolateral pterygoid surface. Thus, the gain, particularly in relation to bars and fenestrae, have led to a question at issue in relation to the lepidosaurian lower temporal variety of hypotheses concerning skull use and kinesis. Of these, one bar is not the functional advantage of its loss (13), but rather of of the most enduring relates to the absence of the lower temporal bar its gain in some rhynchocephalians and, very rarely, in lizards in squamates and the acquisition of streptostyly. We performed a (12, 14). This, in turn, raises questions as to the selective series of computer modeling studies on the skull of Uromastyx advantages of the different lepidosaurian skull morphologies. hardwickii, an akinetic herbivorous lizard. Multibody dynamic anal- Morphological changes in the lepidosaurian skull have been ysis (MDA) was conducted to predict the forces acting on the skull, the subject of theoretical and experimental studies (6, 8, 9, and the results were transferred to a finite element analysis (FEA) to 15–19) that aimed to understand the underlying selective crite- estimate the pattern of stress distribution. -

Lacertoid Footprints from the Upper Triassic (Santa Maria Formation) of Southern Brazil
Palaeogeography, Palaeoclimatology, Palaeoecology 262 (2008) 140–156 Contents lists available at ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo Lacertoid footprints from the Upper Triassic (Santa Maria Formation) of Southern Brazil Rafael Costa da Silva a,⁎, Jorge Ferigolo b, Ismar de Souza Carvalho a, Antonio Carlos Sequeira Fernandes c a CCMN, IGEO, Departamento de Geologia, Universidade Federal do Rio de Janeiro, Cidade Universitária, Ilha do Fundão, Av. Athos da Silveira Ramos, 274, Bloco G, 21910-200, Rio de Janeiro, RJ, Brazil b Museu de Ciências Naturais, Fundação Zoobotânica do Rio Grande do Sul, Av. Dr. S. França, 1427, 90690-000, Porto Alegre, RS, Brazil c Museu Nacional/UFRJ, Quinta da Boa Vista, São Cristóvão, 20940-040, Rio de Janeiro, RJ, Brazil article info abstract Article history: The Triassic of Southern Brazil is well-known for its rich tetrapod body fossils. However, trace fossils such as Received 12 January 2007 tetrapod trackways and trails were discovered only recently from the Predebon outcrop (upper part of the Received in revised form 31 January 2008 Alemoa Member, Santa Maria Formation), providing new information on small-sized animals. The trackways Accepted 18 February 2008 can be identified as Rhynchosauroides retroversipes isp. nov., Rhynchosauroides isp., Rhynchosauroides? isp. and drag marks produced by swimming animals of lacertoid affinity. The preservation of the tracks and Keywords: trackways was influenced by the water level whilst they were being made and subsequent subaerial exposure. Rhynchosauroides retroversipes isp. nov. The trackmaker of R. retroversipes isp. nov. corresponds to a lacertoid quadruped tetrapod with primitive Lacertoid footprints autopodia, a sprawling gait and possibly a long tail, whilst the pes was directed postero-laterally.