Saprolegniaceae
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Mass Flow in Hyphae of the Oomycete Achlya Bisexualis
Mass flow in hyphae of the oomycete Achlya bisexualis A thesis submitted in partial fulfilment of the requirements for the Degree of Master of Science in Cellular and Molecular Biology in the University of Canterbury by Mona Bidanjiri University of Canterbury 2018 Abstract Oomycetes and fungi grow in a polarized manner through the process of tip growth. This is a complex process, involving extension at the apex of the cell and the movement of the cytoplasm forward, as the tip extends. The mechanisms that underlie this growth are not clearly understood, but it is thought that the process is driven by the tip yielding to turgor pressure. Mass flow, the process where bulk flow of material occurs down a pressure gradient, may play a role in tip growth moving the cytoplasm forward. This has previously been demonstrated in mycelia of the oomycete Achlya bisexualis and in single hypha of the fungus Neurospora crassa. Microinjected silicone oil droplets were observed to move in the predicted direction after the establishment of an imposed pressure gradient. In order to test for mass flow in a single hypha of A. bisexualis the work in this thesis describes the microinjection of silicone oil droplets into hyphae. Pressure gradients were imposed by the addition of hyperosmotic and hypoosmotic solutions to the hyphae. In majority of experiments, after both hypo- and hyperosmotic treatments, the oil droplets moved down the imposed gradient in the predicted direction. This supports the existence of mass flow in single hypha of A. bisexualis. The Hagen-Poiseuille equation was used to calculate the theoretical rate of mass flow occurring within the hypha and this was compared to observed rates. -

Molecular Identification of Fungi
Molecular Identification of Fungi Youssuf Gherbawy l Kerstin Voigt Editors Molecular Identification of Fungi Editors Prof. Dr. Youssuf Gherbawy Dr. Kerstin Voigt South Valley University University of Jena Faculty of Science School of Biology and Pharmacy Department of Botany Institute of Microbiology 83523 Qena, Egypt Neugasse 25 [email protected] 07743 Jena, Germany [email protected] ISBN 978-3-642-05041-1 e-ISBN 978-3-642-05042-8 DOI 10.1007/978-3-642-05042-8 Springer Heidelberg Dordrecht London New York Library of Congress Control Number: 2009938949 # Springer-Verlag Berlin Heidelberg 2010 This work is subject to copyright. All rights are reserved, whether the whole or part of the material is concerned, specifically the rights of translation, reprinting, reuse of illustrations, recitation, broadcasting, reproduction on microfilm or in any other way, and storage in data banks. Duplication of this publication or parts thereof is permitted only under the provisions of the German Copyright Law of September 9, 1965, in its current version, and permission for use must always be obtained from Springer. Violations are liable to prosecution under the German Copyright Law. The use of general descriptive names, registered names, trademarks, etc. in this publication does not imply, even in the absence of a specific statement, that such names are exempt from the relevant protective laws and regulations and therefore free for general use. Cover design: WMXDesign GmbH, Heidelberg, Germany, kindly supported by ‘leopardy.com’ Printed on acid-free paper Springer is part of Springer Science+Business Media (www.springer.com) Dedicated to Prof. Lajos Ferenczy (1930–2004) microbiologist, mycologist and member of the Hungarian Academy of Sciences, one of the most outstanding Hungarian biologists of the twentieth century Preface Fungi comprise a vast variety of microorganisms and are numerically among the most abundant eukaryotes on Earth’s biosphere. -

Saprolegnia Species in Norwegian Salmon Hatcheries: Field Survey Identifies S
Vol. 114: 189–198, 2015 DISEASES OF AQUATIC ORGANISMS Published June 3 doi: 10.3354/dao02863 Dis Aquat Org OPENPEN ACCESSCCESS Saprolegnia species in Norwegian salmon hatcheries: field survey identifies S. diclina sub-clade IIIB as the dominating taxon E. Thoen1, T. Vrålstad1, E. Rolén1, R. Kristensen1, Ø. Evensen2, I. Skaar1,* 1Norwegian Veterinary Institute, PO Box 750 Sentrum, 0106 Oslo, Norway 2Norwegian University of Life Sciences, PO Box 8146 Dep., 0033 Oslo, Norway ABSTRACT: Saprolegnia isolates within the recognized clades encompassing the taxa S. parasit- ica and S. diclina act as opportunist and aggressive pathogens to both fish and their eggs. They are responsible for significant economic losses in aquaculture, particularly in salmonid hatcheries. However, the identity, distribution and pathogenic significance of involved species often remain unexplored. In this study, 89 Saprolegnia isolates were recovered from water, eggs and salmon tis- sue samples that originated from salmon (Salmo salar) hatcheries along the coast of Norway. The cultures were characterized morphologically and molecularly in order to provide an overview of the species composition of Saprolegnia spp. present in Norwegian salmon hatcheries. We demon- strate that S. diclina clearly dominated and contributed to 79% of the recovered isolates. Parsi- mony analyses of the nuclear ribosomal internal transcribed spacer (ITS) region split these isolates into 2 strongly supported sub-clades, S. diclina sub-clade IIIA and IIIB, where sub-clade IIIB accounted for 66% of all isolates. A minor portion of the isolates constituted other taxa that were either conspecific or showed strong affinity to S. parasitica, S. ferax, S. hypogyna and Scoliolegnia asterophora. -

The Taxonomy and Biology of Phytophthora and Pythium
Journal of Bacteriology & Mycology: Open Access Review Article Open Access The taxonomy and biology of Phytophthora and Pythium Abstract Volume 6 Issue 1 - 2018 The genera Phytophthora and Pythium include many economically important species Hon H Ho which have been placed in Kingdom Chromista or Kingdom Straminipila, distinct from Department of Biology, State University of New York, USA Kingdom Fungi. Their taxonomic problems, basic biology and economic importance have been reviewed. Morphologically, both genera are very similar in having coenocytic, hyaline Correspondence: Hon H Ho, Professor of Biology, State and freely branching mycelia, oogonia with usually single oospores but the definitive University of New York, New Paltz, NY 12561, USA, differentiation between them lies in the mode of zoospore differentiation and discharge. Email [email protected] In Phytophthora, the zoospores are differentiated within the sporangium proper and when mature, released in an evanescent vesicle at the sporangial apex, whereas in Pythium, the Received: January 23, 2018 | Published: February 12, 2018 protoplast of a sporangium is transferred usually through an exit tube to a thin vesicle outside the sporangium where zoospores are differentiated and released upon the rupture of the vesicle. Many species of Phytophthora are destructive pathogens of especially dicotyledonous woody trees, shrubs and herbaceous plants whereas Pythium species attacked primarily monocotyledonous herbaceous plants, whereas some cause diseases in fishes, red algae and mammals including humans. However, several mycoparasitic and entomopathogenic species of Pythium have been utilized respectively, to successfully control other plant pathogenic fungi and harmful insects including mosquitoes while the others utilized to produce valuable chemicals for pharmacy and food industry. -

Ajb205620.Pdf
AMERICAN JOURNAL OF BOTANY VOL. VIII MAY, 1921 NO·5 ISOACHLYA, A NEW GENUS OF THE SAPROLEGNIACEAEI C. H. KAUFFMAN (Received for publication December 29, 1920) Isoachlya Kauffman gen. nov. Hyphae rather stout or slender. Zoo sporangia formed from their tips, oval, pyriform, ventricose-clavate, the later ones (secondary) arising either by cymose or pseudo-cymose arrange ment as in Achlya, or by internal proliferation as in Saprolegnia, both modes occuring earlier or later in the development of one and the same species, or frequently on the same main hypha. Zoospores diplanetic, as in Saproleg nia, escaping and swarming separately, and after encystment swarming the second time before the formation of a germ tube. Oogonia terminal or toru lose, occasionally intercalary. Oospores with centric contents, the spores filling the oogonium incompletely. Antheridia present or few to none. The genus is characterized and distinguished, in the main, by the presence of the cymose or Achlya mode of formation of secondary sporangia, coupled with diplanetic zoospores. The following species naturally fall within its boundaries: I. Isoachlya toruloides Kauffman and Coker sp. nov. 2. Isoachlya paradoxa (Coker) comb. nov. Achlya paradoxa Coker. Mycologia 6: 285. 1914. 3. Isoachlya monilifera (de Bary) comb. nov. Saprolegnia monilifera de Bary. Bot. Zeit. 16: 629. 1888. Isoachlya toruloides Kauffman and Coker sp. nov. Hyphae rather slender and short, 18-20" in diameter, later ones fre quently smaller, straight and scarcely branched. Zoosporangia oval, pyriform, clavate-pyriform, more rarely elongated-pyriform, with a more or less distinct papilla; secondary sporangia, during the early and vigorous develdpment, all cymosely arranged by successive basipetal formation, sometimes from the walls of earlier ones, later secondary sporangial initials appearing by internal proliferation as in Saprolegnia; zoospores diplanetic, capable of escaping and swarming separately, encysting after coming to 1 After this paper was in the hands of the editor, a letter from Prof. -

Differentiation and Pathogenicity Within the Saprolegniaceae
Comprehensive Summaries of Uppsala Dissertations from the Faculty of Science and Technology 680 _____________________________ _____________________________ Differentiation and Pathogenicity within the Saprolegniaceae Studies on Physiology and Gene Expression Patterns in Saprolegnia parasitica and Aphanomyces astaci BY GUNNAR ANDERSSON ACTA UNIVERSITATIS UPSALIENSIS UPPSALA 2001 Dissertation for the Degree of Doctor of Philosophy in Physiological Mycology presented at Uppsala University in 2002 Abstract Andersson, M. G. 2001. Differentiation and Pathogenicity within the Saprolegniaceae. Studies on Physiology and Gene Expression Patterns in Saprolegnia parasitica and Aphanomyces astaci. Acta Universitatis Upsaliensis. Comprehensive Summaries of Uppsala Dissertations from the Faculty of Science and Technology 680, 41 pp. Uppsala. ISBN 91-554-5203-5. Saprolegnia parasitica and Aphanomyces astaci are parasitic water moulds belonging to the Oomycetes. Despite their importance as parasites they are very little studied at the molecular level and the work described in this thesis was aimed at increasing the molecular knowledge of these organisms by cloning and characterising genes of potential importance for reproduction and pathogenicity. Stage-specific transcripts from Saprolegnia parasitica were isolated by differential display RT-PCR. One of the markers, puf1 encodes a putative mRNA binding protein which may be involved in post-transcriptional regulation of gene expression. S. parasitica puf1 is expressed exclusively in spore cysts that have not been determined for germination or repeated zoospore emergence indicating that the cyst stage has two phases, of about equal duration, which are physiologically and transcriptionally distinct. A similar expression pattern is observed in Aphanomyces spp. with different regulation of spore development and in the transcript is detected in both primary and secondary cysts. -

The Classification of Lower Organisms
The Classification of Lower Organisms Ernst Hkinrich Haickei, in 1874 From Rolschc (1906). By permission of Macrae Smith Company. C f3 The Classification of LOWER ORGANISMS By HERBERT FAULKNER COPELAND \ PACIFIC ^.,^,kfi^..^ BOOKS PALO ALTO, CALIFORNIA Copyright 1956 by Herbert F. Copeland Library of Congress Catalog Card Number 56-7944 Published by PACIFIC BOOKS Palo Alto, California Printed and bound in the United States of America CONTENTS Chapter Page I. Introduction 1 II. An Essay on Nomenclature 6 III. Kingdom Mychota 12 Phylum Archezoa 17 Class 1. Schizophyta 18 Order 1. Schizosporea 18 Order 2. Actinomycetalea 24 Order 3. Caulobacterialea 25 Class 2. Myxoschizomycetes 27 Order 1. Myxobactralea 27 Order 2. Spirochaetalea 28 Class 3. Archiplastidea 29 Order 1. Rhodobacteria 31 Order 2. Sphaerotilalea 33 Order 3. Coccogonea 33 Order 4. Gloiophycea 33 IV. Kingdom Protoctista 37 V. Phylum Rhodophyta 40 Class 1. Bangialea 41 Order Bangiacea 41 Class 2. Heterocarpea 44 Order 1. Cryptospermea 47 Order 2. Sphaerococcoidea 47 Order 3. Gelidialea 49 Order 4. Furccllariea 50 Order 5. Coeloblastea 51 Order 6. Floridea 51 VI. Phylum Phaeophyta 53 Class 1. Heterokonta 55 Order 1. Ochromonadalea 57 Order 2. Silicoflagellata 61 Order 3. Vaucheriacea 63 Order 4. Choanoflagellata 67 Order 5. Hyphochytrialea 69 Class 2. Bacillariacea 69 Order 1. Disciformia 73 Order 2. Diatomea 74 Class 3. Oomycetes 76 Order 1. Saprolegnina 77 Order 2. Peronosporina 80 Order 3. Lagenidialea 81 Class 4. Melanophycea 82 Order 1 . Phaeozoosporea 86 Order 2. Sphacelarialea 86 Order 3. Dictyotea 86 Order 4. Sporochnoidea 87 V ly Chapter Page Orders. Cutlerialea 88 Order 6. -

Connecticut Aquatic Nuisance Species Management Plan
CONNECTICUT AQUATIC NUISANCE SPECIES MANAGEMENT PLAN Connecticut Aquatic Nuisance Species Working Group TABLE OF CONTENTS Table of Contents 3 Acknowledgements 5 Executive Summary 6 1. INTRODUCTION 10 1.1. Scope of the ANS Problem in Connecticut 10 1.2. Relationship with other ANS Plans 10 1.3. The Development of the CT ANS Plan (Process and Participants) 11 1.3.1. The CT ANS Sub-Committees 11 1.3.2. Scientific Review Process 12 1.3.3. Public Review Process 12 1.3.4. Agency Review Process 12 2. PROBLEM DEFINITION AND RANKING 13 2.1. History and Biogeography of ANS in CT 13 2.2. Current and Potential Impacts of ANS in CT 15 2.2.1. Economic Impacts 16 2.2.2. Biodiversity and Ecosystem Impacts 19 2.3. Priority Aquatic Nuisance Species 19 2.3.1. Established ANS Priority Species or Species Groups 21 2.3.2. Potentially Threatening ANS Priority Species or Species Groups 23 2.4. Priority Vectors 23 2.5. Priorities for Action 23 3. EXISTING AUTHORITIES AND PROGRAMS 30 3.1. International Authorities and Programs 30 3.2. Federal Authorities and Programs 31 3.3. Regional Authorities and Programs 37 3.4. State Authorities and Programs 39 3.5. Local Authorities and Programs 45 4. GOALS 47 3 5. OBJECTIVES, STRATEGIES, AND ACTIONS 48 6. IMPLEMENTATION TABLE 72 7. PROGRAM MONITORING AND EVALUATION 80 Glossary* 81 Appendix A. Listings of Known Non-Native ANS and Potential ANS in Connecticut 83 Appendix B. Descriptions of Species Identified as ANS or Potential ANS 93 Appendix C. -

Synopsis of Freshwater Crayfish Diseases and Commensal Organisms Brett .F Edgerton James Cook University, [email protected]
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Faculty Publications from the Harold W. Manter Parasitology, Harold W. Manter Laboratory of Laboratory of Parasitology 3-2002 Synopsis of Freshwater Crayfish Diseases and Commensal Organisms Brett .F Edgerton James Cook University, [email protected] Louis H. Evans Curtin University of Technology Frances J. Stephens Curtin University of Technology Robin M. Overstreet Gulf Coast Research Laboratory, [email protected] Follow this and additional works at: https://digitalcommons.unl.edu/parasitologyfacpubs Part of the Aquaculture and Fisheries Commons, and the Parasitology Commons Edgerton, Brett .;F Evans, Louis H.; Stephens, Frances J.; and Overstreet, Robin M., "Synopsis of Freshwater Crayfish Diseases and Commensal Organisms" (2002). Faculty Publications from the Harold W. Manter Laboratory of Parasitology. 884. https://digitalcommons.unl.edu/parasitologyfacpubs/884 This Article is brought to you for free and open access by the Parasitology, Harold W. Manter Laboratory of at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Faculty Publications from the Harold W. Manter Laboratory of Parasitology by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. Published in Aquaculture 206:1–2 (March 2002), pp. 57–135; doi: 10.1016/S0044-8486(01)00865-1 Copyright © 2002 Elsevier Science. Creative Commons Attribution Non-Commercial No Deriva- tives License. Accepted October 18, 2001; published online November 30, 2001. Synopsis of Freshwater Crayfish Diseases and Commensal Organisms Brett F. Edgerton,1 Louis H. Evans,2 Frances J. Stephens,2 and Robin M. Overstreet3 1. Department of Microbiology and Immunology, James Cook University, Townsville, QLD 4810, Australia 2. -

The Effect of Malchite Green As a Fungicide
THE EFFECT OF MALACHITE GREEN AS A FUNGICIDE RICHARD MARTIN Department of Zoology, The Ohio State University, Columbus, Ohio $210 ABSTRACT A series of tests with malachite green, used at various concentrations and treatment durations, were made on six species of watermolds known to be parasitic on fish and/or fish eggs. The tests indicate that these species have markedly different tolerances to the dye. While a treatment of as little as 1 ppm of malachite green for five minutes was effec- tive for Saprolegnia parasitica, it took 10 ppm for five minutes to control Achlya ambi- sexualis and Allomyces macrogynus, and 15 ppm for one hour to control Achlya oblongata. INTRODUCTION The occurrence of parasitic fungi constitutes a chronic problem in fish hatcheries and aquaria. Fish suffering from stress and injury may become infected by fungi, resulting in considerable economic loss, particularly where valuable aquarium and cultured fish are involved. Nigrelli (1943) reported that over 45 deaths in one year were incurred among fishes at the New York Aquarium due to infestations by unidentified species of the genus Saprolegnia. Many fish hatcheries have serious problems with fungal infections of fish and fish eggs (Glen L. Hoffman, 196 X personal communication). The most common treatment for parasitized fish and fish eggs is malachite green oxalate. This aniline dye is used as a dip or flush to eliminate or prevent the growth of fungi. Many studies have been made to try and determine the most effective concentrations for fungicidal-fungistatic effects. Prominent among them is that of O'Donnell (1941), who found that a 10-30-second dip in 67 ppm (1:15,000) malachite green was effective and non-toxic to 18 species of fish. -
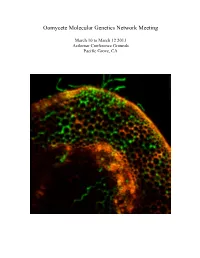
Meeting Program
Oomycete Molecular Genetics Network Meeting March 10 to March 12 2013 Asilomar Conference Grounds Pacific Grove, CA Oomycete Molecular Genetics Network Meeting March 10 to March 12, 2013 Asilomar Conference Grounds Pacific Grove, CA The Oomycete Molecular Genetics Research Network was initially funded by an NSF Research Coordination Network grant in 2001. The purpose of our annual meeting is to promote communication and collaboration, and minimize duplication of effort within the worldwide Oomycete molecular genetics community. The oomycete molecular genetics community now numbers well over one hundred labs from across the world, and papers on oomycetes attract a readership that goes well beyond the community itself. The Oomycete Molecular Genetics Conference alternates between USA and Eurasia, and returns this year to Asilomar, California. This year’s meeting will cover some of the latest research on Effector Biology, Genomics and Oomycete evolution and population biology. With over 100 attendees expected, this meeting represents the largest network meeting held in the US. Committee Chairs John McDowell Mark Gijzen Associate Professor Research Scientist Plant Pathology, Physiology and Weed Science Agriculture and Agri-food Canada Virginia Tech, Blacksburg, VA 24061 London ON, Canada NSV4T3 Meeting Logistics Facilities Coordinator Joel Shuman Paul Morris Project Manager Professor, Biological Sciences Plant Pathology, Physiology and Weed Science Bowling Green State University Virginia Tech, Blacksburg, VA24061 Bowling Green, OH 43403 Cover: Confocal image of GFP-expressing Phytophthora sojae at 12h after inoculation of soybean hypocotyls. Image courtesy of Kai-Tao from the lab of Yuanchao Wang, Nanjing Agricultural University, Nanjing, China. Meeting Sponsors This meeting is supported in part by NSF grant MCB-0639226 to Brett Tyler OMGN 2013 Program Location: Talks will be held at Kiln on Sunday Morning and at Fred Farr for all other sessions. -

Mitochondrial Glycolysis in a Major Lineage of Eukaryotes
GBE Mitochondrial Glycolysis in a Major Lineage of Eukaryotes Carolina Rıo Bartulos 1,2, Matthew B. Rogers3,9, Tom A. Williams4, Eleni Gentekaki5,10, Henner Brinkmann6,11, Ru¨digerCerff1, Marie-Franc¸oise Liaud1,AdrianB.Hehl7, Nigel R. Yarlett8, Ansgar Gruber2,5,12, Peter G. Kroth2,*, and Mark van der Giezen3,* 1Institut fu¨ r Genetik, Technische Universitat€ Braunschweig 2Fachbereich Biologie, Universitat€ Konstanz, Germany 3Biosciences, University of Exeter, United Kingdom 4School of Biological Sciences, University of Bristol, United Kingdom 5Department of Biochemistry & Molecular Biology, Dalhousie University, Halifax, Canada 6Departement de Biochimie, Universite de Montreal C.P. 6128, Montreal, Quebec, Canada 7Institute of Parasitology, University of Zu¨ rich, Switzerland 8Department of Chemistry and Physical Sciences, Pace University 9Present address: Rangos Research Center, University of Pittsburgh, Children’s Hospital, Pittsburgh, PA 10Present address: School of Science and Human Gut Microbiome for Health Research Unit, Mae Fah Luang University, Chiang Rai, Thailand 11Present address: Leibniz-Institut DSMZ—Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Braunschweig, Germany 12Present address: Institute of Parasitology, Biology Centre, Czech Academy of Sciences, Cesk e Budejovice, Czech Republic *Corresponding authors: E-mails: [email protected]; [email protected]. Accepted: July 28, 2018 Abstract The establishment of the mitochondrion is seen as a transformational step in the origin of eukaryotes. With the mitochon- drion came bioenergetic freedom to explore novel evolutionary space leading to the eukaryotic radiation known today. The tight integration of the bacterial endosymbiont with its archaeal host was accompanied by a massive endosymbiotic gene transfer resulting in a small mitochondrial genome which is just a ghost of the original incoming bacterial genome.