Baleen’ Whale Rsos.Royalsocietypublishing.Org (Mysticeti: Aetiocetidae)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Arktocara Yakataga, a New Fossil Odontocete (Mammalia, Cetacea) from the Oligocene of Alaska and the Antiquity of Platanistoidea
Arktocara yakataga, a new fossil odontocete (Mammalia, Cetacea) from the Oligocene of Alaska and the antiquity of Platanistoidea Alexandra T. Boersma1,2 and Nicholas D. Pyenson1,3 1 Department of Paleobiology, National Museum of Natural History, Smithsonian Institution, Washington, D.C., United States of America 2 College of Extended Education, California State University, Monterey Bay, Seaside, CA, United States of America 3 Departments of Mammology and Paleontology, Burke Museum of Natural History and Culture, Seattle, WA, United States of America ABSTRACT The diversification of crown cetacean lineages (i.e., crown Odontoceti and crown Mysticeti) occurred throughout the Oligocene, but it remains an ongoing challenge to resolve the phylogenetic pattern of their origins, especially with respect to stem lineages. One extant monotypic lineage, Platanista gangetica (the Ganges and Indus river dolphin), is the sole surviving member of the broader group Platanistoidea, with many fossil relatives that range from Oligocene to Miocene in age. Curiously, the highly threatened Platanista is restricted today to freshwater river systems of South Asia, yet nearly all fossil platanistoids are known globally from marine rocks, suggesting a marine ancestry for this group. In recent years, studies on the phylogenetic relationships in Platanistoidea have reached a general consensus about the membership of different sub-clades and putative extinct groups, although the position of some platanistoid groups (e.g., Waipatiidae) has been contested. Here we describe a new genus and species of fossil platanistoid, Arktocara yakataga, gen. et sp. nov. from the Oligocene of Alaska, USA. The type and only known specimen was collected from the marine Poul Submitted 16 May 2016 Creek Formation, a unit known to include Oligocene strata, exposed in the Yakutat Accepted 13 July 2016 City and Borough of Southeast Alaska. -

PDF of Manuscript and Figures
The triple origin of whales DAVID PETERS Independent Researcher 311 Collinsville Avenue, Collinsville, IL 62234, USA [email protected] 314-323-7776 July 13, 2018 RH: PETERS—TRIPLE ORIGIN OF WHALES Keywords: Cetacea, Mysticeti, Odontoceti, Phylogenetic analyis ABSTRACT—Workers presume the traditional whale clade, Cetacea, is monophyletic when they support a hypothesis of relationships for baleen whales (Mysticeti) rooted on stem members of the toothed whale clade (Odontoceti). Here a wider gamut phylogenetic analysis recovers Archaeoceti + Odontoceti far apart from Mysticeti and right whales apart from other mysticetes. The three whale clades had semi-aquatic ancestors with four limbs. The clade Odontoceti arises from a lineage that includes archaeocetids, pakicetids, tenrecs, elephant shrews and anagalids: all predators. The clade Mysticeti arises from a lineage that includes desmostylians, anthracobunids, cambaytheres, hippos and mesonychids: none predators. Right whales are derived from a sister to Desmostylus. Other mysticetes arise from a sister to the RBCM specimen attributed to Behemotops. Basal mysticetes include Caperea (for right whales) and Miocaperea (for all other mysticetes). Cetotheres are not related to aetiocetids. Whales and hippos are not related to artiodactyls. Rather the artiodactyl-type ankle found in basal archaeocetes is also found in the tenrec/odontocete clade. Former mesonychids, Sinonyx and Andrewsarchus, nest close to tenrecs. These are novel observations and hypotheses of mammal interrelationships based on morphology and a wide gamut taxon list that includes relevant taxa that prior studies ignored. Here some taxa are tested together for the first time, so they nest together for the first time. INTRODUCTION Marx and Fordyce (2015) reported the genesis of the baleen whale clade (Mysticeti) extended back to Zygorhiza, Physeter and other toothed whales (Archaeoceti + Odontoceti). -

From Peru Sheds Light on the Early Miocene Platanistoid Disparity And
A new large squalodelphinid (Cetacea, Odontoceti) from Peru sheds light on the Early Miocene platanistoid disparity and ecology Giovanni Bianucci, Giulia Bosio, Elisa Malinverno, Christian de Muizon, Igor Villa, Mario Urbina, Olivier Lambert To cite this version: Giovanni Bianucci, Giulia Bosio, Elisa Malinverno, Christian de Muizon, Igor Villa, et al.. A new large squalodelphinid (Cetacea, Odontoceti) from Peru sheds light on the Early Miocene platanis- toid disparity and ecology. Royal Society Open Science, The Royal Society, 2018, 5 (4), pp.172302. 10.1098/rsos.172302. hal-02611968 HAL Id: hal-02611968 https://hal.archives-ouvertes.fr/hal-02611968 Submitted on 18 May 2020 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Downloaded from http://rsos.royalsocietypublishing.org/ on April 18, 2018 A new large squalodelphinid (Cetacea, rsos.royalsocietypublishing.org Odontoceti) from Peru Research sheds light on the Early Cite this article: Bianucci G, Bosio G, Miocene platanistoid Malinverno E, de Muizon C, Villa IM, Urbina M, Lambert O. 2018 A new large squalodelphinid (Cetacea, Odontoceti) from Peru sheds light on disparity and ecology the Early Miocene platanistoid disparity and 1 2 2 ecology. R. Soc. -

How to Cite Complete Issue More Information About This Article
Boletín de la Sociedad Geológica Mexicana ISSN: 1405-3322 Sociedad Geológica Mexicana, A.C. Hernández Cisneros, Atzcalli Ehécatl; González Barba, Gerardo; Fordyce, Robert Ewan Oligocene cetaceans from Baja California Sur, Mexico Boletín de la Sociedad Geológica Mexicana, vol. 69, no. 1, January-April, 2017, pp. 149-173 Sociedad Geológica Mexicana, A.C. Available in: http://www.redalyc.org/articulo.oa?id=94350664007 How to cite Complete issue Scientific Information System Redalyc More information about this article Network of Scientific Journals from Latin America and the Caribbean, Spain and Portugal Journal's homepage in redalyc.org Project academic non-profit, developed under the open access initiative Boletín de la Sociedad Geológica Mexicana / 2017 / 149 Oligocene cetaceans from Baja California Sur, Mexico Atzcalli Ehécatl Hernández Cisneros, Gerardo González Barba, Robert Ewan Fordyce ABSTRACT Atzcalli Ehecatl Hernández Cisneros ABSTRACT RESUMEN [email protected] Museo de Historia Natural de la Universidad Autónoma de Baja California Sur, Univer- Baja California Sur has an import- Baja California Sur tiene un importante re- sidad Autónoma de Baja California Sur, ant Cenozoic marine fossil record gistro de fósiles marinos del Cenozoico que Carretera al Sur Km 5.5, Apartado Postal which includes diverse but poorly incluye los restos poco conocidos de cetáceos 19-B, C.P. 23080, La Paz, Baja California Sur, México. known Oligocene cetaceans from del Oligoceno de México. En este estudio Instituto Politécnico Nacional, Centro Inter- Mexico. Here we review the cetacean ofrecemos más detalles sobre estos fósiles de disciplinario de Ciencias Marinas (CICMAR), fossil record including new observa- cetáceos, incluyendo nuevas observaciones Av. Instituto Politécnico Nacional s/n, Col. -

The Biology of Marine Mammals
Romero, A. 2009. The Biology of Marine Mammals. The Biology of Marine Mammals Aldemaro Romero, Ph.D. Arkansas State University Jonesboro, AR 2009 2 INTRODUCTION Dear students, 3 Chapter 1 Introduction to Marine Mammals 1.1. Overture Humans have always been fascinated with marine mammals. These creatures have been the basis of mythical tales since Antiquity. For centuries naturalists classified them as fish. Today they are symbols of the environmental movement as well as the source of heated controversies: whether we are dealing with the clubbing pub seals in the Arctic or whaling by industrialized nations, marine mammals continue to be a hot issue in science, politics, economics, and ethics. But if we want to better understand these issues, we need to learn more about marine mammal biology. The problem is that, despite increased research efforts, only in the last two decades we have made significant progress in learning about these creatures. And yet, that knowledge is largely limited to a handful of species because they are either relatively easy to observe in nature or because they can be studied in captivity. Still, because of television documentaries, ‘coffee-table’ books, displays in many aquaria around the world, and a growing whale and dolphin watching industry, people believe that they have a certain familiarity with many species of marine mammals (for more on the relationship between humans and marine mammals such as whales, see Ellis 1991, Forestell 2002). As late as 2002, a new species of beaked whale was being reported (Delbout et al. 2002), in 2003 a new species of baleen whale was described (Wada et al. -

Evolution of River Dolphins
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Publications, Agencies and Staff of the U.S. Department of Commerce U.S. Department of Commerce 3-7-2001 Evolution of river dolphins Healy Hamilton Susana Caballero Allen G. Collins Robert L. Brownell Jr. NOAA, [email protected] Follow this and additional works at: https://digitalcommons.unl.edu/usdeptcommercepub Part of the Environmental Sciences Commons Hamilton, Healy; Caballero, Susana; Collins, Allen G.; and Brownell, Robert L. Jr., "Evolution of river dolphins" (2001). Publications, Agencies and Staff of the U.S. Department of Commerce. 111. https://digitalcommons.unl.edu/usdeptcommercepub/111 This Article is brought to you for free and open access by the U.S. Department of Commerce at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Publications, Agencies and Staff of the U.S. Department of Commerce by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. Proceedings: Biological Sciences, Vol. 268, No. 1466 (Mar. 7, 2001), pp. 549-556 B THE ROYAL doi 10.1098/rspb.2000.1385 92 SOCIETY Evolution of river dolphins Healy E{amiltonltsSusana Caballero2,Allen G. Coll;nsl and Robert L. BrownellJr3 lMuseumof Paleontologyand Department of IntegrativeBiology, University of California,Berkeley, CA 94720, USA 2FundacionSubarta, Carrara 24Foeste, no 3-110,Cali, Colombia 3SouthwestFisheries Science Center, PO Box 271,La jrolla, CA 92038, USA The world's river dolphins (Inia, Pontoporia,Lipotes and Platanista)are among the least known and most endangered of all cetaceans. The four extant genera inhabit geographically disjunct river systems and exhibit highly modified morphologies, leading many cetologists to regard river dolphins as an unnatural group. -

Gigantism Precedes Filter Feeding in Baleen Whale Evolution
1 This is the preprint version of the following article: Fordyce, R.E., and Marx, F.G. (2018). Gigantism 2 precedes filter feeding in baleen whale evolution. Curr Biol 28, 1670-1676.e1672, which has been 3 published in final form at https://www.cell.com/current-biology/fulltext/S0960-9822(18)30455- 4 X?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS096098221830455X 5 %3Fshowall%3Dtrue 6 7 Gigantism precedes filter feeding in baleen whale evolution 8 R. Ewan Fordyce1,2 and Felix G. Marx3-5* 9 1Department of Geology, University of Otago, Dunedin, New Zealand 10 2Departments of Vertebrate Zoology and Paleobiology, National Museum of Natural History, Smithsonian Institution, 11 Washington DC, USA 12 5Directorate of Earth and History of Life, Royal Belgian Institute of Natural Sciences, Brussels, Belgium 13 3School of Biological Sciences, Monash University, Clayton, Victoria, Australia 14 4Geosciences, Museum Victoria, Melbourne, Australia 15 *E-mail: [email protected] 16 17 SUMMARY 18 Baleen whales (Mysticeti) are the largest animals on Earth, thanks to their ability to filter feed vast 19 amounts of small prey from seawater. Whales appeared during the latest Eocene, but evidence of 20 their early evolution remains both sparse and controversial [1, 2], with several models competing 21 to explain the origin of baleen-based bulk feeding [3-6]. Here, we describe a virtually complete 22 skull of Llanocetus denticrenatus, the second-oldest (ca 34 Ma) mysticete known. The new 23 material represents the same individual as the type and only specimen, a fragmentary mandible. 24 Phylogenetic analysis groups Llanocetus with the oldest mysticete, Mystacodon selenensis [2], 25 into the basal family Llanocetidae. -

New Discoveries of Fossil Toothed Whales from Peru: Our Changing Perspective of Beaked Whale and Sperm Whale Evolution
Quad. Mus. St. Nat. Livorno, 23: 13-27 (2011) DOI code: 10.4457/musmed.2010.23.13 13 New discoveries of fossil toothed whales from Peru: our changing perspective of beaked whale and sperm whale evolution OLIVIER LAMBERT1 SUMMARY: Following the preliminary description of a first fossil odontocete (toothed whale) from the Miocene of the Pisco Formation, southern coast of Peru, in 1944, many new taxa from Miocene and Pliocene levels of this formation were described during the 80’s and 90’s, (families Kentriodontidae, Odobenocetopsidae, Phocoenidae, and Pontoporiidae). Only one Pliocene Ziphiidae (beaked whale) and one late Miocene Kogiidae (dwarf sperm whale) were defined. Modern beaked whales and sperm whales (Physeteroidea = Kogiidae + Physeteridae) share several ecological features: most are predominantly teuthophagous, suction feeders, and deep divers. They further display a highly modified cranial and mandibular morphology, including tooth reduction in both groups, high vertex and sexually dimorphic mandibular tusks in ziphiids, and development of a vast supracranial basin in physeteroids. New discoveries from the Miocene of the Pisco Formation enrich the fossil record of ziphiids and physeteroids and shed light on various aspects of their evolution. From Cerro Colorado, a new species of the ziphiid Messapicetus lead to the description of features previously unknown in fossil members of the family: association of complete upper and lower tooth series with tusks, hypothetical sexual dimorphism in the development of the tusks, skull anatomy of a calf... A new small ziphiid from Cerro los Quesos, Nazcacetus urbinai, is characterized by the reduction of the dentition: a pair of apical mandibular tusks associated to vestigial postapical teeth, likely hold in the gum. -

This Article Is Essentially a Hoax for April 1 of 2014 For
NOTE: THIS ARTICLE IS ESSENTIALLY A HOAX FOR APRIL 1 OF 2014 FOR ASHORO MUSEUM OF PALEONTOLOGY, BUT AIMS TO BE A THOUGHT-PROVOKING PIECE OF WORK TO CONSIDER THE BODY SIZE EVOLUTION IN MYSTICETI (BALEEN WHALES). A Lilliputian whale, Aetiocetus hobbitus Tsai, Cheng-Hsiu Department of Geology, University of Otago, Dunedin, New Zealand Whales (Cetacea: Mysticeti) are the largest animals in the world. As the real Brobdingnagians, how small a whale could be renders an interesting and unresolved biological question. Here I report a smallest whale “produced” at the Ashoro Museum of Paleontology (AMP), Hokkaido, Japan. SYSTEMATIC PALAEONTOLOGY Cetacea Brisson, 1762 Mysticeti Gray, 1864 Aetiocetidae Emlong, 1966 Aetiocetus Emlong, 1966 Aetiocetus hobbitus sp. nov. (FIG. 1) Holotype: AMP0401, including a well-preserved skull and left mandible. Diagnosis: The morphology of AMP0401 is “exactly” the same as Aetiocetus polydentatus, but the size is considerably smaller. The skull length is about 15 cm. Etymology: The species epithet, hobbitus, is latinised from The Hobbit written by J. R. R. Tolkien, describing a fictional, diminutive human species. Aetiocetus hobbitus represents the smallest mysticete species described and named to date. Body size evolution in baleen whales is particularly tantalising and intriguing, since baleen whales are the largest animals to have ever lived on Earth. Given the body size of the earliest known mysticete, Llanocetus denticrenatus, the body size has evolved to both directions in Mysticeti, gigantism: Balaenoptera musculus (blue whale) and dwarfism: Aetiocetus hobbitus sp. nov. (this study). Understanding the underlying evolutionary process that influences the body size evolution in baleen whales in at least two different directions should help decipher the mystery of mysteries – from Lilliputian to Brobdingnagian in baleen whale evolution. -
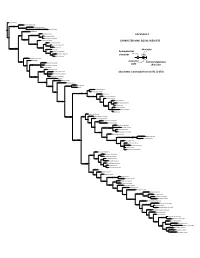
Appendices 2-6 As PDF for Download
Zygorhiza kochii 19 61 128148160 Archaeodelphis patrius 11 13 42 44 62 64 105159161 1 1 1 1 1 39 43 45 52 102150220 3 2 1 1 1 1 1 2 2 20 50 81 86 88 96 165 Waipatia maerewhenua 1 1 1 1 1 1 2 9 17 19 36 49 56 77 89 100103106107116120133145146151162163171196203210 1 2 1 1 1 1 4 Physeter catodon 1 3 2 1 2 1 1 1 1 1 1 1 2 1 1 1 1 1 3 1 1 2 1 2 27 41 46 102113148155188 ChM PV4745 0 1 1 1 1 1 1 1 23 26 Janjucetus hunderi 85 152 4 22 24 40 66 67 71 126213220 1 0 APPENDIX 2 1 1 2 1 1 2 0 1 1 0 1 2 34 188 Mammalodon colliveri 1 1 Mammalodon hakataramea 0 32 39 42 154161 101136 0 1 1 1 1 2 Morawanocetus yabukii 1 1 CHARACTER MAP, EQUAL WEIGHTS 71 136 Chonecetus sookensis 2 18 69 89 98 99 190228 100 1 1 1 1 1 1 1 1 1 2 0 110111 113 Chonecetus goedertorum 69 148238 28 33 78 220242 2 3 1 Fucaia buelli 3 38 54 57 76 100126134252 0 1 1 0 1 1 3 1 42 82 107111 145 1 1 1 1 1 1 1 1 1 OCPC 1178 0 0 1 1 2 character 97 0 0 Aetiocetus polydentatus 60 101106 1 homoplasious 73 126201 0 1 1 136 Aetiocetus cotylalveus 1 0 0 188 character 1 Aetiocetus weltoni 158 3 35 46 48 61 99 Llanocetus denticrenatus 0 1 1 1 1 4 39 71 OU GS10897 2 18 72 138 0 1 1 1 90 98 99 103129 character 1 1 1 1 Sitsqwayk cornisorum non homoplasious 0 1 1 1 2 119135145167170180181183184186187 109137268 13 113 state Tlaxcallicetus guaycurae character 5 1 3 1 1 1 1 1 2 1 1 1 1 1 1 1 205 Tlaxcallicetus sp. -

How Modern Baleen Whales Arose from a Fossil “Dark Age”
Editors' choice Like phoenix from the ashes: How modern baleen whales arose from a fossil “dark age” FELIX G. MARX, ERICH M.G. FITZGERALD, and R. EWAN FORDYCE Marx, F.G., Fitzgerald, E.M.G., and Fordyce, R.E. 2019. Like phoenix from the ashes: How modern baleen whales arose from a fossil “dark age”. Acta Palaeontologica Polonica 64 (2): 231–238. The evolution of baleen whales (Mysticeti), the largest animals on Earth, was punctuated by a pivotal turnover event. Following their emergence around 36 million years (Ma), mysticetes diversified into a disparate range of toothed and toothless species until 23 Ma, but then nearly vanished from the global fossil record for the next five million years. Following this early Miocene “dark age”, toothless mysticetes spectacularly reappeared around 18–17 Ma, whereas toothed mysticetes had gone entirely extinct. Here, we suggest that this turnover event reflects a change in mysticete habitat occupancy. Using the well-sampled record of Australasia as a case study, we show that Oligocene pre-“dark age” mysticetes formed distinct coastal and offshore assemblages, dominated by small (2–4 m), ecologically disparate toothed species, and larger (5–6 m) toothless filter feeders, respectively. Environmental change around the Oligocene–Miocene boundary led to the decline of the endemic coastal assemblages, leaving nearshore deposits virtually devoid of mys- ticetes. Filter feeders persisted offshore and subsequently re-invaded coastal habitats during the mid-Miocene Climatic Optimum, thus establishing the modern, cosmopolitan mysticete fauna. Key words: Mammalia, Mysticeti, evolution, Oligocene, Miocene, Zealandia, Australia. Felix G. Marx [[email protected]], Directorate Earth and History of Life, Royal Belgian Institute of Natural Sciences, Rue Vautier 29, Brussels 1000, Belgium; Department of Geology, University of Liège, Belgium; School of Biological Sciences, Monash University, Clayton, Vic., Australia; Museums Victoria, Melbourne, Vic., Australia. -

Mysticetes Baring Their Teeth: a New Fossil Whale, Mammalodon Hakataramea, from the Southwest Pacific
Memoirs of Museum Victoria 74: 107–116 (2016) Published 2016 ISSN 1447-2546 (Print) 1447-2554 (On-line) http://museumvictoria.com.au/about/books-and-journals/journals/memoirs-of-museum-victoria/ Mysticetes baring their teeth: a new fossil whale, Mammalodon hakataramea, from the Southwest Pacific R. EWAN FORDYCE1,2,* (http://zoobank.org/urn:lsid:zoobank.org:author:311048BF-4642-412E-B5DA-E01B8C03B802) AND FELIX G. MARX1,3 (http://zoobank.org/urn:lsid:zoobank.org:author:1791C478-33A7-4C75-8104-4C98C7B22125) 1 Department of Geology, University of Otago, PO Box 56, Dunedin 9054, New Zealand ([email protected]) 2 Departments of Vertebrate Zoology and Paleobiology, National Museum of Natural History, Smithsonian Institution, Washington DC 20560, USA 3 Department of Geology and Palaeontology, National Museum of Nature and Science, Tsukuba, Japan (felix.marx@ otago.ac.nz) * To whom correspondence should be addressed. E-mail: [email protected] http://zoobank.org/urn:lsid:zoobank.org:pub:7A2CAF55-70DC-4561-AA3D-86FA72C721E6 Abstract Fordyce, R.E. and Marx, F.G. 2016. Mysticetes baring their teeth: a new fossil whale, Mammalodon hakataramea, from the Southwest Pacific. Memoirs of Museum Victoria 74: 107–116. A small, toothed fossil cetacean from Hakataramea Valley (South Canterbury, New Zealand) represents a new Late Oligocene species, Mammalodon hakataramea. The new material is from the Kokoamu Greensand (Duntroonian Stage, about 27 Ma, early to middle Chattian) of the Canterbury Basin, and thus about 2 Ma older than the only other species included in this genus, Mammalodon colliveri (Late Oligocene, Victoria, Australia). The anterior pedicle of the tympanic bulla is not fused to the periotic and resembles that of Delphinidae in basic structure.