MAPPING PLANT SPECIES RANGES the Hawaiian ISLANDS
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Handbook Publication.Pub
Table of Contents Maui County’s Landscape and Gardening Handbook Xeriscaping in Maui County ................................................................. 1 Planning and Design................................................................................................................. 1 Hydro-zones.............................................................................................................................. 1 Plant Selection and the Maui jkCounty Planting Zones............................................................ 2 Soil Preparation ........................................................................................................................ 4 Mulching.................................................................................................................................... 5 Irrigation .................................................................................................................................... 5 Maintenance ............................................................................................................................. 7 Other Interesting Techniques for the Ambitious ..................................... 8 Xeriscape Ponds....................................................................................................................... 8 Aquaponics in the Backyard ..................................................................................................... 9 Water Polymer Crystals ........................................................................................................... -

Life in Mauna Kea's Alpine Desert
Life in Mauna Kea’s by Mike Richardson Alpine Desert Lycosa wolf spiders, a centipede that preys on moribund insects that are blown to the summit, and the unique, flightless wekiu bug. A candidate for federal listing as an endangered species, the wekiu bug was first discovered in 1979 by entomologists on Pu‘u Wekiu, the summit cinder cone. “Wekiu” is Hawaiian for “topmost” or “summit.” The wekiu bug belongs to the family Lygaeidae within the order of insects known as Heteroptera (true bugs). Most of the 26 endemic Hawaiian Nysius species use a tube-like beak to feed on native plant seed heads, but the wekiu bug uses its beak to suck the hemolymph (blood) from other insects. High above the sunny beaches, rocky coastline, Excluding its close relative Nysius a‘a on the nearby Mauna Loa, the wekiu bug and lush, tropical forests of the Big Island of Hawai‘i differs from all the world’s 106 Nysius lies a unique environment unknown even to many species in its predatory habits and unusual physical characteristics. The bug residents. The harsh, barren, cold alpine desert is so possesses nearly microscopically small hostile that it may appear devoid of life. However, a wings and has the longest, thinnest legs few species existing nowhere else have formed a pre- and the most elongated head of any Lygaeid bug in the world. carious ecosystem-in-miniature of insects, spiders, The wekiu bug, about the size of a other arthropods, and simple plants and lichens. Wel grain of rice, is most often found under rocks and cinders where it preys diur come to the summit of Mauna Kea! nally (during daylight) on insects and Rising 13,796 feet (4,205 meters) dependent) community of arthropods even birds that are blown up from lower above sea level, Mauna Kea is the was uncovered at the summit. -
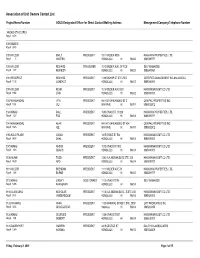
Association of Unit Owners Contact List
Association of Unit Owners Contact List Project Name/Number AOUO Designated Officer for Direct Contact/Mailing Address Management Company/Telephone Number `AKOKO AT HO`OPILI Reg.# 8073 1001 QUEEN Reg.# 7675 1001 WILDER EMILY PRESIDENT 1001 WILDER #305 HAWAIIAN PROPERTIES, LTD. Reg.# 5 WATERS HONOLULU HI 96822 8085399777 1010 WILDER RICHARD TREASURER 1010 WILDER AVE, OFFICE SELF MANAGED Reg.# 377 KENNEDY HONOLULU HI 96822 8085241961 1011 PROSPECT RICHARD PRESIDENT 1188 BISHOP ST STE 2503 CERTIFIED MANAGEMENT INC dba ASSOCI Reg.# 1130 CONRADT HONOLULU HI 96813 8088360911 1015 WILDER KEVIN PRESIDENT 1015 WILDER AVE #201 HAWAIIANA MGMT CO LTD Reg.# 1960 LIMA HONOLULU HI 96822 8085939100 1037 KAHUAMOKU VITA PRESIDENT 94-1037 KAHUAMOKU ST 3 CEN PAC PROPERTIES INC Reg.# 1551 VILI WAIPAHU HI 96797 8085932902 1040 KINAU PAUL PRESIDENT 1040 KINAU ST., #1206 HAWAIIAN PROPERTIES, LTD. Reg.# 527 FOX HONOLULU HI 96814 8085399777 1041 KAHUAMOKU ALAN PRESIDENT 94-1041 KAHUAMOKU ST 404 CEN PAC PROPERTIES INC Reg.# 1623 IGE WAIPAHU HI 96797 8085932902 1054 KALO PLACE JUANA PRESIDENT 1415 S KING ST 504 HAWAIIANA MGMT CO LTD Reg.# 5450 DAHL HONOLULU HI 96814 8085939100 1073 KINAU ANSON PRESIDENT 1073 KINAU ST 1003 HAWAIIANA MGMT CO LTD Reg.# 616 QUACH HONOLULU HI 96814 8085939100 1108 AUAHI TODD PRESIDENT 1240 ALA MOANA BLVD STE. 200 HAWAIIANA MGMT CO LTD Reg.# 7429 APO HONOLULU HI 96814 8085939100 1111 WILDER BRENDAN PRESIDENT 1111 WILDER AVE 7A HAWAIIAN PROPERTIES, LTD. Reg.# 228 BURNS HONOLULU HI 96822 8085399777 1112 KINAU LINDA Y SOLE OWNER 1112 KINAU ST PH SELF MANAGED Reg.# 1295 NAKAGAWA HONOLULU HI 96814 1118 ALA MOANA NICHOLAS PRESIDENT 1118 ALA MOANA BLVD., SUITE 200 HAWAIIANA MGMT CO LTD Reg.# 7431 VANDERBOOM HONOLULU HI 96814 8085939100 1133 WAIMANU ANNA PRESIDENT 1133 WAIMANU STREET, STE. -

Mauna Loa Reconnaissance 2003
“Giant of the Pacific” Mauna Loa Reconnaissance 2003 Plan of encampment on Mauna Loa summit illustrated by C. Wilkes, Engraved by N. Gimbrede (Wilkes 1845; vol. IV:155) Prepared by Dennis Dougherty B.A., Project Director Edited by J. Moniz-Nakamura, Ph. D. Principal Investigator Pacific Island Cluster Publications in Anthropology #4 National Park Service Hawai‘i Volcanoes National Park Department of the Interior 2004 “Giant of the Pacific” Mauna Loa Reconnaissance 2003 Prepared by Dennis Dougherty, B.A. Edited by J. Moniz-Nakamura, Ph.D. National Park Service Hawai‘i Volcanoes National Park P.O. Box 52 Hawaii National Park, HI 96718 November, 2004 Mauna Loa Reconnaissance 2003 Executive Summary and Acknowledgements The Mauna Loa Reconnaissance project was designed to generate archival and inventory/survey level recordation for previously known and unknown cultural resources within the high elevation zones (montane, sub-alpine, and alpine) of Mauna Loa. Field survey efforts included collecting GPS data at sites, preparing detailed site plan maps and feature descriptions, providing site assessment and National Register eligibility, and integrating the collected data into existing site data bases within the CRM Division at Hawaii Volcanoes National Park (HAVO). Project implementation included both pedestrian transects and aerial transects to accomplish field survey components and included both NPS and Research Corporation University of Hawaii (RCUH) personnel. Reconnaissance of remote alpine areas was needed to increase existing data on historic and archeological sites on Mauna Loa to allow park managers to better plan for future projects. The reconnaissance report includes a project introduction; background sections including physical descriptions, cultural setting overview, and previous archeological studies; fieldwork sections describing methods, results, and feature and site summaries; and a section on conclusions and findings that provide site significance assessments and recommendations. -

Topographic History of the Maui Nui Complex, Hawai'i, and Its Implications for Biogeography1
Topographic History ofthe Maui Nui Complex, Hawai'i, and Its Implications for Biogeography 1 Jonathan Paul Price 2,4 and Deborah Elliott-Fisk3 Abstract: The Maui Nui complex of the Hawaiian Islands consists of the islands of Maui, Moloka'i, Lana'i, and Kaho'olawe, which were connected as a single landmass in the past. Aspects of volcanic landform construction, island subsi dence, and erosion were modeled to reconstruct the physical history of this complex. This model estimates the timing, duration, and topographic attributes of different island configurations by accounting for volcano growth and subsi dence, changes in sea level, and geomorphological processes. The model indi cates that Maui Nui was a single landmass that reached its maximum areal extent around 1.2 Ma, when it was larger than the current island of Hawai'i. As subsi dence ensued, the island divided during high sea stands of interglacial periods starting around 0.6 Ma; however during lower sea stands of glacial periods, islands reunited. The net effect is that the Maui Nui complex was a single large landmass for more than 75% of its history and included a high proportion of lowland area compared with the contemporary landscape. Because the Hawaiian Archipelago is an isolated system where most of the biota is a result of in situ evolution, landscape history is an important detertninant of biogeographic pat terns. Maui Nui's historical landscape contrasts sharply with the current land scape but is equally relevant to biogeographical analyses. THE HAWAIIAN ISLANDS present an ideal logic histories that can be reconstructed more setting in which to weigh the relative influ easily and accurately than in most regions. -

Invasive Aphids Attack Native Hawaiian Plants
Biol Invasions DOI 10.1007/s10530-006-9045-1 INVASION NOTE Invasive aphids attack native Hawaiian plants Russell H. Messing Æ Michelle N. Tremblay Æ Edward B. Mondor Æ Robert G. Foottit Æ Keith S. Pike Received: 17 July 2006 / Accepted: 25 July 2006 Ó Springer Science+Business Media B.V. 2006 Abstract Invasive species have had devastating plants. To date, aphids have been observed impacts on the fauna and flora of the Hawaiian feeding and reproducing on 64 native Hawaiian Islands. While the negative effects of some inva- plants (16 indigenous species and 48 endemic sive species are obvious, other species are less species) in 32 families. As the majority of these visible, though no less important. Aphids (Ho- plants are endangered, invasive aphids may have moptera: Aphididae) are not native to Hawai’i profound impacts on the island flora. To help but have thoroughly invaded the Island chain, protect unique island ecosystems, we propose that largely as a result of anthropogenic influences. As border vigilance be enhanced to prevent the aphids cause both direct plant feeding damage incursion of new aphids, and that biological con- and transmit numerous pathogenic viruses, it is trol efforts be renewed to mitigate the impact of important to document aphid distributions and existing species. ranges throughout the archipelago. On the basis of an extensive survey of aphid diversity on the Keywords Aphid Æ Aphididae Æ Hawai’i Æ five largest Hawaiian Islands (Hawai’i, Kaua’i, Indigenous plants Æ Invasive species Æ Endemic O’ahu, Maui, and Moloka’i), we provide the first plants Æ Hawaiian Islands Æ Virus evidence that invasive aphids feed not just on agricultural crops, but also on native Hawaiian Introduction R. -

U-Series Isotopic Constraints on Mantle Upwelling on the Periphery of the Hawaiian Plume Isotope Geochemistry of Haleakala Crater Basanites Erin H
U-series isotopic constraints on mantle upwelling on the periphery of the Hawaiian plume Isotope geochemistry of Haleakala crater basanites Erin H. Phillips and Kenneth W.W. Sims, University of Wyoming, and Vincent J.M. Salters, Florida State University 1. Introduction 2. Major and Trace Element Geochemistry and Long-lived Radiogenic Isotopes 100 400 ● The Hawaiian islands are part of the age progressive series of 16 Comparison to Haleakala Crater (this study) 90 global alkaline volcanoes that form the Hawaiian-Emperor seamount chain. At 14 Haleakala SW Rift Zone (Sims et al., 1999) 300 suites Haleakala, on the island of Maui, basanitic lavas of the Hana Haleakala post-shield 80 12 Haleakala shield 200 Volcanics represent end-member rejuvenated stage alkaline 70 Kilauea magmatism (Clauge, 1987). Although shield-stage tholeiitic 100 10 Mauna Loa 60 volcanism predominates in the Hawaiian islands, alkaline lavas O 2 Mauna Kea 0 erupted on the trailing edge of the Hawaiian plume present an 8 Hualalai trachytes 50 O+Na important component to understanding mantle melting and solid 2 Loihi K 40 6 Nyiragongo mantle upwelling. Averages for 30 Leucite Hills lamproites Hawaiian Lavas 4 Ross Island, Antarctica basanites 20 ● We present geochemical data for 13 samples from Haleakala Samoa crater. 14C ages for seven samples range from 870 ± 40 to 4070 ± 50 2 See supplement for full reference list 10 years (Sherrod and McGeehin, 1999). Preliminary data for 5 0 1995) Mantle (McDonough and Sun, Sample/Primitive 0 samples from the Haleakala southwest rift zone (Sims et al., 1999) 35 40 45 50 55 60 65 Rb Ba Th U Nb La Ce Sr Pb Nd Sm Hf Eu Dy Y Yb Lu SiO are consistent with iso-viscous (Watson and McKenzie, 1991) and 2 thermo-viscous (Hauri et al., 1994) fluid mechanical models of 10 plume upwelling in which upwelling rates are slower on the Hawaiian Lavas periphery of plume. -

Hibiscus Arnottianus Subsp. Immaculatus (Koki‘O Ke‘Oke‘O) Current Classification: Endangered
5-YEAR REVIEW Short Form Summary Species Reviewed: Hibiscus arnottianus subsp. immaculatus (koki‘o ke‘oke‘o) Current Classification: Endangered Federal Register Notice announcing initiation of this review: [USFWS] U.S. Fish and Wildlife Service. 2009. Endangered and threatened wildlife and plants; initiation of 5-year reviews of 103 species in Hawaii. Federal Register 74(49):11130-11133. Lead Region/Field Office: Region 1/Pacific Islands Fish and Wildlife Office (PIFWO), Honolulu, Hawaii Name of Reviewer(s): Marie Bruegmann, Plant Recovery Coordinator, PIFWO Jess Newton, Recovery Program Lead, PIFWO Assistant Field Supervisor for Endangered Species, PIFWO Methodology used to complete this 5-year review: This review was conducted by staff of the Pacific Islands Fish and Wildlife Office of the U.S. Fish and Wildlife Service (USFWS), beginning on March 16, 2009. The review was based on final critical habitat designations for Hibiscus arnottianus subsp. immaculatus and other species from the island of Molokai (USFWS 2003) as well as a review of current, available information. The National Tropical Botanical Garden provided an initial draft of portions of the review and recommendations for conservation actions needed prior to the next five-year review. The evaluation of Samuel Aruch, biological consultant, was reviewed by the Plant Recovery Coordinator. The document was then reviewed by the Recovery Program Lead and the Assistant Field Supervisor for Endangered Species before submission to the Field Supervisor for approval. Background: For information regarding the species listing history and other facts, please refer to the Fish and Wildlife Service’s Environmental Conservation On-line System (ECOS) database for threatened and endangered species (http://ecos.fws.gov/tess_public). -

The Fern Family Blechnaceae: Old and New
ANDRÉ LUÍS DE GASPER THE FERN FAMILY BLECHNACEAE: OLD AND NEW GENERA RE-EVALUATED, USING MOLECULAR DATA Tese apresentada ao Programa de Pós-Graduação em Biologia Vegetal do Departamento de Botânica do Instituto de Ciências Biológicas da Universidade Federal de Minas Gerais, como requisito parcial à obtenção do título de Doutor em Biologia Vegetal. Área de Concentração Taxonomia vegetal BELO HORIZONTE – MG 2016 ANDRÉ LUÍS DE GASPER THE FERN FAMILY BLECHNACEAE: OLD AND NEW GENERA RE-EVALUATED, USING MOLECULAR DATA Tese apresentada ao Programa de Pós-Graduação em Biologia Vegetal do Departamento de Botânica do Instituto de Ciências Biológicas da Universidade Federal de Minas Gerais, como requisito parcial à obtenção do título de Doutor em Biologia Vegetal. Área de Concentração Taxonomia Vegetal Orientador: Prof. Dr. Alexandre Salino Universidade Federal de Minas Gerais Coorientador: Prof. Dr. Vinícius Antonio de Oliveira Dittrich Universidade Federal de Juiz de Fora BELO HORIZONTE – MG 2016 Gasper, André Luís. 043 Thefern family blechnaceae : old and new genera re- evaluated, using molecular data [manuscrito] / André Luís Gasper. – 2016. 160 f. : il. ; 29,5 cm. Orientador: Alexandre Salino. Co-orientador: Vinícius Antonio de Oliveira Dittrich. Tese (doutorado) – Universidade Federal de Minas Gerais, Departamento de Botânica. 1. Filogenia - Teses. 2. Samambaia – Teses. 3. RbcL. 4. Rps4. 5. Trnl. 5. TrnF. 6. Biologia vegetal - Teses. I. Salino, Alexandre. II. Dittrich, Vinícius Antônio de Oliveira. III. Universidade Federal de Minas Gerais. Departamento de Botânica. IV. Título. À Sabrina, meus pais e a vida, que não se contém! À Lucia Sevegnani, que não pode ver esta obra concluída, mas que sempre foi motivo de inspiração. -

THE HAWAIIAN-EMPEROR VOLCANIC CHAIN Part I Geologic Evolution
VOLCANISM IN HAWAII Chapter 1 - .-............,. THE HAWAIIAN-EMPEROR VOLCANIC CHAIN Part I Geologic Evolution By David A. Clague and G. Brent Dalrymple ABSTRACT chain, the near-fixity of the hot spot, the chemistry and timing of The Hawaiian-Emperor volcanic chain stretches nearly the eruptions from individual volcanoes, and the detailed geom 6,000 km across the North Pacific Ocean and consists of at least etry of volcanism. None of the geophysical hypotheses pro t 07 individual volcanoes with a total volume of about 1 million posed to date are fully satisfactory. However, the existence of km3• The chain is age progressive with still-active volcanoes at the Hawaiian ewell suggests that hot spots are indeed hot. In the southeast end and 80-75-Ma volcanoes at the northwest addition, both geophysical and geochemical hypotheses suggest end. The bend between the Hawaiian and .Emperor Chains that primitive undegassed mantle material ascends beneath reflects a major change in Pacific plate motion at 43.1 ± 1.4 Ma Hawaii. Petrologic models suggest that this primitive material and probably was caused by collision of the Indian subcontinent reacts with the ocean lithosphere to produce the compositional into Eurasia and the resulting reorganization of oceanic spread range of Hawaiian lava. ing centers and initiation of subduction zones in the western Pacific. The volcanoes of the chain were erupted onto the floor of the Pacific Ocean without regard for the age or preexisting INTRODUCTION structure of the ocean crust. Hawaiian volcanoes erupt lava of distinct chemical com The Hawaiian Islands; the seamounts, hanks, and islands of positions during four major stages in their evolution and the Hawaiian Ridge; and the chain of Emperor Seamounts form an growth. -

Hawaiian Hotspot
V51B-0546 (AGU Fall Meeting) High precision Pb, Sr, and Nd isotope geochemistry of alkalic early Kilauea magmas from the submarine Hilina bench region, and the nature of the Hilina/Kea mantle component J-I Kimura*, TW Sisson**, N Nakano*, ML Coombs**, PW Lipman** *Department of Geoscience, Shimane University **Volcano Hazard Team, USGS JAMSTEC Shinkai 6500 81 65 Emperor Seamounts 56 42 38 27 20 12 5 0 Ma Hawaii Islands Ready to go! Hawaiian Hotspot N ' 0 3 ° 9 Haleakala 1 Kea trend Kohala Loa 5t0r6 end Mauna Kea te eii 95 ol Hualalai Th l 504 na io sit Mauna Loa Kilauea an 509 Tr Papau 208 Hilina Seamont Loihi lic ka 209 Al 505 N 98 ' 207 0 0 508 ° 93 91 9 1 98 KAIKO 99 SHINKAI 02 KAIKO Loa-type tholeiite basalt 209 Transitional basalt 95 504 208 Alkalic basalt 91 508 0 50 km 155°00'W 154°30'W Fig. 1: Samples obtained by Shinkai 6500 and Kaiko dives in 1998-2002 10 T-Alk Mauna Kea 8 Kilauea Basanite Mauna Loa -K M Hilina (low-Si tholeiite) 6 Hilina (transitional) Hilina (alkali) Kea Hilina (basanite) 4 Loa Hilina (nephelinite) Papau (Loa type tholeiite) primary 2 Nep Basaltic Basalt andesite (a) 0 35 40 45 50 55 60 SiO2 Fig. 2: TAS classification and comparison of the early Kilauea lavas to representative lavas from Hawaii Big Island. Kea-type tholeiite through transitional, alkali, basanite to nephelinite lavas were collected from Hilina bench. Loa-type tholeiite from Papau seamount 100 (a) (b) (c) OIB OIB OIB M P / e l p 10 m a S E-MORB E-MORB E-MORB Loa type tholeiite Low Si tholeiite Transitional 1 f f r r r r r r r f d y r r d b b -

Winter 2011 - 1 President’S Message
THE HARDY FERN FOUNDATION P.O. Box 3797 Federal Way, WA 98063-3797 Web site: www.hardvfems.org The Hardy Fern Foundation was founded in 1989 to establish a comprehen¬ sive collection of the world’s hardy ferns for display, testing, evaluation, public education and introduction to the gardening and horticultural community. Many rare and unusual species, hybrids and varieties are being propagated from spores and tested in selected environments for their different degrees of hardiness and ornamental garden value. The primary fern display and test garden is located at, and in conjunction with, The Rhododendron Species Botanical Garden at the Weyerhaeuser Corporate Headquarters, in Federal Way, Washington. Satellite fern gardens are at the Birmingham Botanical Gardens, Birmingham, Alabama, California State University at Sacramento, California, Coastal Maine Botanical Garden, Boothbay , Maine. Dallas Arboretum, Dallas, Texas, Denver Botanic Gardens, Denver, Colorado, Georgeson Botanical Garden, University of Alaska, Fairbanks, Alaska, Harry R Leu Garden, Orlando, Florida, Inniswood Metro Gardens, Columbus, Ohio, New York Botanical Garden, Bronx, New York, and Strybing Arboretum, San Francisco, California. The fern display gardens are at Bainbridge Island Library. Bainbridge Island, WA, Bellevue Botanical Garden, Bellevue, WA, Lakewold, Tacoma, Washington, Lotusland, Santa Barbara, California, Les Jardins de Metis, Quebec, Canada, Rotary Gardens, Janesville, Wl, and Whitehall Historic Home and Garden, Louisville, KY. Hardy Fern Foundation members participate in a spore exchange, receive a quarterly newsletter and have first access to ferns as they are ready for distribution. Cover design by Will anna Bradner HARDY FERN FOUNDATION QUARTERLY THE HARDY FERN FOUNDATION QUARTERLY Volume 21 Editor- Sue Olsen ISSN 154-5517 President’s Message Patrick Kennar American Fern Society - 2010 Fern Foray Tom Stuart Thelypteris noveboracensis Tapering fern, New York fern.