Appalachian Biogeography Symposium
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Wilderness Visitors and Recreation Impacts: Baseline Data Available for Twentieth Century Conditions
United States Department of Agriculture Wilderness Visitors and Forest Service Recreation Impacts: Baseline Rocky Mountain Research Station Data Available for Twentieth General Technical Report RMRS-GTR-117 Century Conditions September 2003 David N. Cole Vita Wright Abstract __________________________________________ Cole, David N.; Wright, Vita. 2003. Wilderness visitors and recreation impacts: baseline data available for twentieth century conditions. Gen. Tech. Rep. RMRS-GTR-117. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 52 p. This report provides an assessment and compilation of recreation-related monitoring data sources across the National Wilderness Preservation System (NWPS). Telephone interviews with managers of all units of the NWPS and a literature search were conducted to locate studies that provide campsite impact data, trail impact data, and information about visitor characteristics. Of the 628 wildernesses that comprised the NWPS in January 2000, 51 percent had baseline campsite data, 9 percent had trail condition data and 24 percent had data on visitor characteristics. Wildernesses managed by the Forest Service and National Park Service were much more likely to have data than wildernesses managed by the Bureau of Land Management and Fish and Wildlife Service. Both unpublished data collected by the management agencies and data published in reports are included. Extensive appendices provide detailed information about available data for every study that we located. These have been organized by wilderness so that it is easy to locate all the information available for each wilderness in the NWPS. Keywords: campsite condition, monitoring, National Wilderness Preservation System, trail condition, visitor characteristics The Authors _______________________________________ David N. -

Shining Rock and Grassy Cove Top Hike
Old Butt Knob Trail and Shining Creek Trail Loop - Shining Rock Wilderness, Pisgah National Forest, NC Length Difficulty Streams Views Solitude Camping 11.9 mls Hiking Time: 8 hours with 2 hours of breaks Elev. Gain: 3,410 ft Parking: Park at the Big East Fork Trailhead on U.S. 276. 35.36583, -82.81786 By Trail Contributor: Zach Robbins The Old Butt Knob Trail and Shining Creek Trail loop is a classic introduction to the Shining Rock Wilderness Area of North Carolina. Beginning at the Big East Fork Trailhead on U.S. 276, both trails climb from 3,384 feet to meet the Art Loeb Trail above 5,800 feet at Shining Rock Gap. Despite the relatively short length of this loop (9.4 miles including Shining Rock), this is a difficult day hike for hikers of all abilities. The Old Butt Knob Trail climbs over 1,400 feet in the first mile, and the Shining Creek Trail is incredibly rocky and steep over its last mile. Despite the hardship, this is an excellent backpacking loop with outstanding campsites and even better views. The Old Butt Knob Trail features multiple views from southern-facing rock outcrops, and the views from Shining Rock and Grassy Cove Top are some of the highlights of the wilderness. Even though this is a wilderness area, this is in close proximity to Asheville and is popular with weekend backpackers. Try to start early if you want to camp at Shining Rock Gap, which is one of the best campsites in the region and is also the crossroads for 4 trails within the wilderness. -

Pisgah District Trails
PISGAH RANGER DISTRICT TRAILS Table of Contents: Trail Name Length Rating Trail Use Page # Andy Cove Nature Trail 0.7mi Easy Hiking 3 Art Loeb Spur 0.6mi Difficult Hiking 4 Art Loeb Trail–Section 1 12.3mi Difficult Hiking 5 Art Loeb Trail-Section 2 7.2mi Difficult Hiking 6 Art Loeb Trail-Section 3 6.8mi Difficult Hiking 7 Art Loeb Trail-Section 4 3.8mi Difficult Hiking 8 Avery Creek 3.2mi Medium Hiking/Biking 9 Bad Fork 2.0mi Difficult Hiking 10 Bennett Gap 2.9mi Medium Hiking/Biking(seasonal) 11 Big Creek 4.9mi Difficult Hiking/Sections open to bikes& horses 12 Biltmore Campus 0.9mi Easy Hiking/ wheelchair accessible 13 Black Mountain 9.8mi Difficult Hiking/Biking 14 Boyd Branch 0.7mi Easy Hiking/Biking 15 Bradley Creek 5.1mi Medium Hiking/Biking/Horses 16 Buck Spring 6.2mi Medium Hiking 17 Buckeye Gap 3.1mi Difficult Hiking 18 Buckhorn Gap 5.2mi Medium Hiking/Biking/Horses 19 Buckwheat Knob 1.5mi Medium Hiking/Biking 20 Butter Gap 3.4mi Medium Hiking/Biking 21 Caney Bottom 4.6mi Medium Hiking/Sections open to biking 22 Case Camp Ridge 1.7mi Difficult Hiking 23 Cat Gap Bypass 0.4mi Easy Hiking 24 Cat Gap Loop 4.4mi Medium Hiking/Sections open to biking(seasonal) 25 Chestnut Cove 0.2mi Medium Hiking 26 Clawhammer Cove 1.5mi Medium Hiking 27 Club Gap 0.8mi Medium Hiking/Biking 28 Cold Mountain 1.4mi Difficult Hiking 29 Coontree Loop 3.7mi Medium Hiking/Sections open to biking(seasonal) 30 Courthouse Falls 0.3mi Easy Hiking 31 Cove Creek 0.7mi Easy Hiking/Biking 32 Daniel Ridge Loop 4.0mi Medium Hiking/Biking 33 Deer Lake Lodge 1.4mi Medium -

Cold Mountain
Cold Mountain Rd. (SR 1129) and proceed 4 The hike starts at the northern miles until it deadends into end of the Art Loeb Trail. At Camp Daniel Boone. Once Deep Gap, it heads north on the Cold Mountain Trail to in the camp stay to the left, the top of the mountain. On drive past the buildings and this hike, you will travel from onto the forest road, and park 3,200 ft. elev. at the trailhead on the right side of the road. to 6,030 ft. at the top, through The trail starts at the Forest several zones of vegetation. Service sign on the left. Your hike up is like going back to an earlier season. Type of hike: Out and back In spring, flowers past their Distance: 9.8 miles prime at lower elevations are still waiting to bloom up top. Total ascent: 3,500 ft. In early autumn, trees at the Highlights: Views, flowers, summit have already changed literary reference, SB6K in color, while foliage is still green the Wilderness Area at the trailhead. Since this hike is in a USGS map: Waynesville, Wilderness Area, the trails are Cruso, NC not blazed. However, because of the increased fame of Cold Trail map: Pisgah Ranger Mountain brought on by the District, Pisgah National book and movie, there is now Forest, National Geographic a wand and sign at the Forest Trails Illustrated #780 Service information board. Land managed by: Pisgah National Forest, Pisgah District Related book & movie: Cold Getting to the trailhead: Mountain by Charles Frazier From Waynesville, take US and the 2003 film of the same 276 south for 6.5 miles. -

Kevin Adams 250 North Carolina Waterfalls Challenge
Kevin Adams 250 North Carolina Waterfalls Challenge Trail LengthTrail Hike Difficulty Page # Number Waterfall Name Hub Date Viewed Notes 1-100 Kevin Adams 100 NC Waterfalls Challenge 101 Window Falls Hanging Rock & Mayo River 0.6 5-7 27 102 Upper Cascade Falls Hanging Rock & Mayo River 0.3 2-5 28 103 Lower Cascade Falls Hanging Rock & Mayo River 0.4 3-7 29 104 Fall Creek Falls Hanging Rock & Mayo River 300 yards 2 31 105 Stone Mountain Falls Stone Mountain State Park 0.6 3-5 34 106 Cascade Falls Boone 0.4 3 39 107 Glen Burney Falls Blowing Rock 1.07 6 42 108 Glen Marie Falls Blowing Rock 1.2 6 42 109 Thunderhole Falls Blowing Rock 0.5 5 44 110 Hebron Colony Falls Blowing Rock 1.6 3-6 45 111 Raven Cliff Falls Grandfather Mountain 2.5 10 48 112 Burnthouse Branch Falls Grandfather Mountain 3 8 51 113 Upper Gragg Prong Falls Grandfather Mountain 58 114 Harper Creek Falls Grandfather Mountain 1.72 5-8 59 115 Phillips Branch Falls Grandfather Mountain 0.5 4 60 116 Thorps Creek Falls Grandfather Mountain 0.19 3 61 117 Teacups Falls Grandfather Mountain 65 118 Upper Steels Creek Falls Grandfather Mountain 66 119 Otter Falls Grandfather Mountain 69 120 Buckeye Creek Falls Elk Park 0.5 6 75 121 She Falls Falls Elk Park 76 122 Duggers Falls Linville Gorge 0.1 3 80 123 English Falls Linville Gorge 0.4 10 80 124 Still Falls Linville Gorge 1.25 6 83 125 Yellow Fork Falls Linville Gorge 1 5-8 85 126 Cathedral Falls Linville Gorge 2.4 7 86 127 Grassy Creek Falls Little Switzerland 0.9 5 89 128 Cutler Falls Spruce Pine 91 129 Whiteoak Creek Falls Mount -
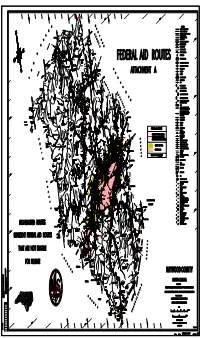
For Release Highlighted Routes
TENN. N.C. ID none 750,000' 780,000' 810,000' 870,000' 840,000' 750,000' 720,000' Rd. 83 00’ 00" 1334 Creek Government M A D I S O N E Wesley LEGEND RD. Springs Creek 82 55’ 00" PRIMITIVE OR UNIMPROVED ROAD Rd. SNOWBIRD E 83 05’ 00" GRADED AND DRAINED ROAD Z Cold 35 45’ 00" Z Snowbird 1334 C O U N T Y SOIL,GRAVEL OR STONE SURFACED ROAD N Great Smoky Mountains S HARD SURFACED ROAD Z 5.9 4 4 4 LANE UNDIVIDED HIGHWAY PEAKED KNOB M O U N T A I N S 5.9 Z Rd. WesleyRd. Creek3.3 3.3 Creek Z Z Rough Knob Peaked Knob S DIVIDED HIGHWAY 690,000' Great Smoky Mountains Harmon Den Wildlife Springs ROUGH KNOB Government Management Area HIGHWAY WITH FRONTAGE ROADS Creek 2.6 Rd. GOV. Patch Harmon Den Wildlife F C A 1334 Max 1334 Management Area HARMON Cold FULL CONTROL ACCESS 2.6 1334 TO NEWPORT Rd. FCA E $40 NS 1336 NON-SYSTEM ROAD FCA Creek Pigeon b 40 DEN 1336 C O U N T Y b $ WILDLIFE PROJECTED LOCATION 1332 1334 Big Wesley F River 1335 F 7.0 F Rd. 1332 40 Sprins .4 F dRd Max R 1335 BelmontRd..2 INTERSECTION DISTANCE PIGEON .2 E 1.3 Church 7.0 E $ 1336 Waterville Rd. Waterville 40 j $ Creek j PUG KNOB 1397 1.2 Creek 6.3 Z Harmon Den Wildlife N Creek Pug Knob Waterville Management Area Creek 1.2 Patch 1397 Harmon Den Wildlife Z TRAFFIC CIRCLE 6.3 Rd. -

Mountains and Meadowlands Along the Blue Ridge Parkway
Mountains and Meadowlands Mountains and Meadowlands Along the Blue Ridge Parkway Text and photos by William A. Bake Office of Publications National Park Service U.S. Department of the Interior Washington, D.C. 1975 A Prologue to the Past 1 The Blue Ridge Parkway Rolling into the distance like "the great distance like breakers and swells Ocean itself," the Blue Ridge dips into a means mountains—the highest deep cove at Wildcat Rocks in Doughton frozen in time. of them mantled with trees and Park (Milepost 240), then swells toward the horizon. Their time, in fact, began long wildflowers uniquely their own. ago, even in geologic terms. The It also means meadows—some Blue Ridge Mountains—indeed, for farming, others that beckon all the Appalachians—are an visitors to wander and explore. cient mountains. Rounded in This booklet introduces both form and subdued in height, the mood and substance of the they are nevertheless a rugged Parkway's highest peaks and lot whose wrinkles and crannies, sweeping meadowlands. Though Indian pipe, a small but conspicuous saphrophyte, pushes past the clover-like cliffs and gorges are often hid designed to be read quickly, it leaves of wood sorrel. den from easy view. remains to be re-examined when ever mountains and meadow- Like so many of the Earth's lands return to mind or presence. mountains, the Appalachians were created in a very complex An "Ocean of Woods swelled manner. It is sufficient to say, and depressed with a waving rather than going through the Surface like that of the great complexities, that what remains Ocean itself wrote an early is a range of mountains which visitor as he sought a way to begins in the Canadian maritime describe the Blue Ridge Moun provinces near the Atlantic and tains. -

Red Crossbill Observations in Western North Carolina MARCUS B
Red Crossbill Observations in Western North Carolina MARCUS B. SIMPSON JR. Department of Pathology, Yale University School of Medicine 310 Cedar Street, New Haven, Connecticut 06510 15 December 1973 The Red Crossbill (Loxia curvirostra) is an uncommon to rare erratic resident through much of the southern Appalachian mountains. The majority of records have come from the Great Smoky Mountains (Stupka, 1963), and observations from other locales are infrequent. The paucity of breeding evidence and the infrequency of large concentrations make the following records significant. SHINING ROCK LEDGE (Haywood County, N.C.) On 13 June 1970, I spent 2 hours watching a pair of Red Crossbills as they constructed a nest in a 30-foot red spruce (Picea rubens) at Shining Spring (elevation 5,740 feet) just S of Shining Rock Gap. During most of the period, the male perched near the top of the spruce and uttered various call notes and songs, while the female carried small twigs to the loosely formed nest, which was located 12 feet from the ground in the mid-canopy of the tree. Whenever I approached the nest, both birds quickly became agitated and gave alarm notes repeatedly. The nest was apparently still in the construction stage, and no eggs or young were found. After leaving the site, I saw flocks of 15 to 20 crossbills at Ivestor Gap (5,700 feet) and at Shining Rock (6,000 feet). Much to my regret, I was unable to return to this isolated locale at a later date to determine whether the breeding had continued. Stupka (1963) and Johnston (1963) have reported evidence for breeding by the Red Crossbill in the southern Appalachians. -

Paul Dickens and the Wilderness Crew
FOURTH QUARTER 2012 Quarterly News Bulletin and Hike Schedule P.O. Box 68, Asheville, NC 28802 • www.carolinamountainclub.org • e-mail: [email protected] Paul Dickens and the Wilderness Crew Becky Smucker and Bill Newton formed a team leader and important cog in through the Middle Prong Wilderness new maintenance crew in the spring of 2008. the machinery. We thought the best in 2007, I hiked other trails in the Since then it has evolved a great deal, and way to update the progress of this Middle Prong that were becoming Paul Dickens has become Becky’s back-up innovative crew was to talk with Paul overgrown and almost impassible Dickens. from years of maintenance neglect. How did you get involved in trail Something had to be done. I start- maintenance and the CMC? ed by recruiting family members to In summer of 2007, my wife and help brush out the Green Mountain I hiked the Big Butt Trail near Mt trail that connected to my section of Mitchell. Parts of that trail were ter- the MST. At about the same time, ribly overgrown with blackberries, Becky Smucker started the Saturday almost impassible. I found the CMC Wilderness Crew to meet two needs website and made a trail problem – to provide an opportunity for CMC report. Shortly afterwards, I received members who work weekdays to do an e-mail from the Asheville Friday trail maintenance on Saturday and Trail Crew asking if I would like to to address the serious need for trail help them clear the blackberry mess. reclamation in the Middle Prong and I took a day off from work, went out Shining Rock Wilderness areas. -

The High Peaks and Asheville—MST Segment 3
1 | Mountains-to-Sea Trail Last Updated 1/1/2017 Approaching Blackstock Knob Photo by Jeff Clark www.internetbrothers.org The High Peaks and Asheville—MST Segment 3 PISGAH INN TO BLACK MOUNTAIN CAMPGROUND By Jim Grode Segment 3 is where Asheville comes to play. It is the most heavily used part of the MST within the mountains, and you are likely to encounter hikers, dog walkers, and trail runners at any time throughout the year. Virtually all of it is easily accessible from the Blue Ridge Parkway (BRP), and there are road crossings every few miles for most of the length. Don’t let its accessibility and popularity fool you, however: With eleva- tions ranging from 2,000 feet at the French Broad River to over 6,600 feet at the summit of Mount Mitchell, this segment is quite rugged. Indeed, the Segment 3 | 2 section just west of Asheville hosts the infamous Shut-In Ridge Trail Run, an 18-mile trail run that annually humbles racers from around the country. Complementing the natural beauty of the Blue Ridge Mountains in this area is the vibrancy of Asheville, a city of 80,000 nestled in the French Broad River valley, which regularly makes lists of the top 10 cities in the United States. Crammed with restaurants, shops, art galleries, and brew- eries, Asheville offers something for nearly everyone and is well worth a layover in your hiking schedule. HIGHLIGHTS INCLUDE • The views atop 6,684-foot Mount Mitchell, the highest point east of the Mississippi River • The Shut-In Trail, which follows the old carriage road from the Biltmore House to George Vanderbilt’s hunting lodge on Mount Pisgah (which no longer stands, but a few remnants of which are still visible) • The cultural and scientific displays at the Blue Ridge Parkway Visitor Center & Headquarters near Asheville • The fine collection of southern art and crafts at the Folk Art Center also near Asheville. -

Fall 2014 Tent
This year’s Tent Peg brings forth a variety of student and faculty experi- ences all brought together into one publication. It is our hope that you as the reader will take these various events and receive the motivation to get out and create your own adventures. Many thanks to the authors and you, the readers! Spencer Williams & Katie Reid 2 Articles Page Debby’s Top 10 Hikes (or Adventures in Hiking) 4 PRM Accomplishments 8 A New Adventure 9 Phased Retirement 10 My Trip to Schoolhouse Falls 11 Who is Pulling the Bowstring Harder? A Look at the True Spiritual Connection of the Outdoors 12 Pinnacle Peak 13 Not A Typical Job For A 20 Year Old 13 A Mountainous Climb 14 The Best Job a Soccer Fan Could Ask For 15 My trip to the Boundary Waters 15 Playing in the Mud 16 Every Fish Is A Blessing: Big Or Small 17 Disc Golf In The Great Smoky Mountains 17 Breaking The Ice 17 Opening Day in Mississippi 18 A Look at the Transformative Power of Wilderness Therapy 19 Sawyer Squeeze Water Filtration System Review 20 Bear lake 20 A Day in the Life of Thomas Graham at Mount Hood, Oregon 21 Strength and Conditioning 21 Adventure Education Conference 22 Special Thanks 23 3 Debby’s Top 10 Hikes (or Adventures in Hiking) 9. Big Creek, Great Smoky Mountains National David Letterman and I have a few things in common. Both Park: (Moderate, various distances) Located on the of our first names begin with the letter “D”. He once worked north side of the GSMNP off of I-40, exit 451 in Tennes- as a weatherman with an off beat humorous take on report- see. -

Peak to Peak—MST Segment 1
1 | Mountains-to-Sea Trail Last Updated 8/5/2019 View from the Mountains-to-Sea Trail at Clingmans Dome Photo by Danny Bernstein Peak to Peak—MST Segment 1 CLINGMANS DOME TO WATERROCK KNOB By Jim Grode The 46.8-mile-long Segment 1 of the Mountains-to-Sea Trail (MST) be- gins in Great Smoky Mountains National Park (GSMNP), straddling Ten- nessee and North Carolina. Newfound Gap Road (US 441), which travels north from Cherokee, North Carolina, to Gatlinburg, Tennessee, forms the backbone of the park and climbs to over 5,000 feet at Newfound Gap. If national parks have specialties, the Smokies is known as a hiker’s park. The scenery is diverse: mountain views, old-growth trees, waterfalls, streams, and more shades of green than a paint chart. Mile for mile, you’ll Segment 1 | 2 find hiking in the park easier than in the neighboring national forests. Even though there are no blazes on Smokies trails, they are so well marked at every intersection that you can follow them with confidence. (Still, stop at a Visitor Center and pick up a Great Smoky Mountains Trail Map for $1.00.) The Smokies may be the most visited national park in the country but only the roads and parking lots are congested. With over 800 miles of trails, even popular trails are not very busy. The trails in the Smokies are well maintained and well marked. Hikers will appreciate that they’re in a na- tional park. The Smokies, in a temperate rainforest, have a great variety of wildflowers, from the first bloodroot in March to the last asters in Octo- ber, as well as more tree species than all of Europe.