Adec Preview Generated PDF File
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Environmental Guidance for Planning and Development
Part A Environmental protection and land use planning in Western Australia Environmental Guidance for Part B Biophysical factors Planning and Development Part C Pollution management May 2008 Part D Social surroundings Guidance Statement No. 33 2007389-0508-50 Foreword The Environmental Protection Authority (EPA) is an independent statutory authority and is the key provider of independent environmental advice to Government. The EPA’s objectives are to protect the environment and to prevent, control and abate pollution and environmental harm. The EPA aims to achieve some of this through the development of environmental protection guidance statements for the environmental impact assessment (EIA) of proposals. This document is one in a series being issued by the EPA to assist proponents, consultants and the public generally to gain additional information about the EPA’s thinking in relation to aspects of the EIA process. The series provides the basis for EPA’s evaluation of, and advice on, proposals under S38 and S48A of the Environmental Protection Act 1986 (EP Act) subject to EIA. The guidance statements are one part of assisting proponents, decision-making authorities and others in achieving environmentally acceptable outcomes. Consistent with the notion of continuous environmental improvement and adaptive environmental management, the EPA expects proponents to take all reasonable and practicable measures to protect the environment and to view the requirements of this Guidance as representing the minimum standards necessary. The main purposes of this EPA guidance statement are: • to provide information and advice to assist participants in land use planning and development processes to protect, conserve and enhance the environment • to describe the processes the EPA may apply under the EP Act to land use planning and development in Western Australia, and in particular to describe the environmental impact assessment (EIA) process applied by the EPA to schemes. -

Annual Report 2010–2011
Australian Museum 2010–2011 Report Annual Australian Museum Annual Report 2010–2011 Australian Museum Annual Report 2010 – 2011 ii Australian Museum Annual Report 2010 –11 The Australian Museum Annual Report 2010 –11 Availability is published by the Australian Museum Trust, This annual report has been designed for accessible 6 College Street Sydney NSW 2010. online use and distribution. A limited number of copies have been printed for statutory purposes. © Australian Museum Trust 2011 This report is available at: ISSN 1039-4141 www.australianmuseum.net.au/Annual-Reports. Editorial Further information on the research and education Project management: Wendy Rapee programs and services of the Australian Museum Editing and typesetting: Brendan Atkins can be found at www.australianmuseum.net.au. Proofreading: Lindsay Taaffe Design and production: Australian Museum Environmental responsibility Design Studio Printed on Sovereign Offset, an FSC- certified paper from responsibly grown fibres, made under an All photographs © Australian Museum ISO 14001– accredited environmental management 2011, unless otherwise indicated. system and without the use of elemental chlorine. Contact Australian Museum 6 College Street Sydney NSW 2010 Open daily 9.30 am – 5.00 pm t 02 9320 6000 f 02 9320 6050 e [email protected] w www.australianmuseum.net.au www.facebook.com/australianmuseum www.twitter.com/austmus www.youtube.com/austmus front cover: The Museum's after-hours program, Jurassic Lounge, attracted a young adult audience to enjoy art, music and new ideas. Photo Stuart Humphreys. iii Minister The Hon. George Souris, MP and Minister for the Arts Governance The Museum is governed by a Trust established under the Australian Museum Trust Act 1975. -

East Kimberley Impact Assessment Project
East Kimberley Impact Assessment Project HISTORICAL NOTES RELEVANT TO IMPACT STORIES OF THE EAST KIMBERLEY Cathie Clement* East Kimberley Working Paper No. 29 ISBN O 86740 357 8 ISSN 0816...,6323 A Joint Project Of The: Centre for Resource and Environmental Studies Australian National University Australian Institute of Aboriginal Studies Anthropology Department University of Western Australia Academy of the Social Sciences in Australia The aims of the project are as follows: 1. To compile a comprehensive profile of the contemporary social environment of the East Kimberley region utilising both existing information sources and limited fieldwork. 2. Develop and utilise appropriate methodological approaches to social impact assessment within a multi-disciplinary framework. 3. Assess the social impact of major public and private developments of the East Kimberley region's resources (physical, mineral and environmental) on resident Aboriginal communities. Attempt to identify problems/issues which, while possibly dormant at present, are likely to have implications that will affect communities at some stage in the future. 4. Establish a framework to allow the dissemination of research results to Aboriginal communities so as to enable them to develop their own strategies for dealing with social impact issues. 5. To identify in consultation with Governments and regional interests issues and problems which may be susceptible to further research. Views expressed in the Projecfs publications are the views of the authors, and are not necessarily shared by the sponsoring organisations. Address correspondence to: The Executive Officer East Kimberley Project CRES, ANU GPO Box4 Canberra City, ACT 2601 HISTORICAL NOTES RELEVANT TO IMPACT STORIES OF THE EAST KIMBERLEY Cathie Clement* East Kimberley Working Paper No. -

BULLETIN of the FLORIDA STATE MUSEUM Biological Sciences
BULLETIN of the FLORIDA STATE MUSEUM Biological Sciences VOLUME 29 1983 NUMBER 3 NON-MARINE MOLLUSKS OF BORNEO II PULMONATA: PUPILLIDAE, CLAUSILIIDAE III PROSOBRANCHIA: HYDROCENIDAE, HELICINIDAE FRED G. THOMPSON AND S. PETER DANCE UNIVERSITY OF FLORIDA GAINESVILLE Numbers of the BULLETIN OF THE FLORIDA STATE MUSEUM, BIOLOGICAL SCIENCES, are published at irregular intervals. Volumes contain about 300 pages and are not necessarily completed in any one calendar year. OLIVER L. AUSTIN, JR., Editor RHODA J . BRYANT, Managing Editor Consultants for this issue: JOHN B. BURCH WILLIAM L. PRATT Communications concerning purchase or exchange of the publications and all manuscripts should be addressed to: Managing Editor, Bulletin; Florida State Museum; University of Florida; Gainesville, FL 32611, U.S.A. Copyright © by the Florida State Museum of the University of Florida This public document was promulgated at an annual cost of $3,040.00 or $3.04 per copy. It makes available to libraries, scholars, and allinterested persons the results of researches in the natural sciences, emphasizing the circum-Caribbean region. Publication dates: 8-15-83 Price: 3.10 NON-MARINE MOLLUSKS OF BORNEO II PULMONATA: PUPILLIDAE, CLAUSILIIDAE III PROSOBRANCHIA: HYDROCENIDAE, HELICINIDAE FRED G. THOMPSON AND S. PETER DANCEl ABSTRACT: The Bornean land snails of the families Pupillidae, Clausiliidae, Hydrocenidae, and Helicinidae are reviewed based on collections from38 localities in Sarawak and Sabah and on previous records from the island. The following species are recorded: PUPILLIDAE- Pupisoma orcula (Benson), Costigo putuiusculum (Issel) new combination, Costigo molecul- ina Benthem-Jutting, Nesopupa moreleti (Brown), N. malayana Issel; Boysidia (Dasypupa) salpimf new subgenus and species, B. -
Explanatory Notes on the Lansdowne Geological Sheet
/52- ally a erric G EOLOG 1 CA 1. SU R V E Y OF W F.STEWN AUSTRAL. I A I)~Re.('loff: J. )-I. LORI) Compiled by D. C. ~e~lat~yand G. errick EX The Lansdowne 1 : 250,000 Sheet area lies in the Kimberley Land Division in the north-eastern corner of Western Australia. It is bounded by longitudes 126" E and 127" 30'E and by latitudes 17" S and 18" S. Lansdowne homestead, near the centre of the Sheet area, is about 300 miles by road from Derby and 340 from Wyndham. These towns are connected by way of Halls Creek by a regularly maintained gravel road. Spasmodically maintained station tracks give access to the Sheet area from Fitzroy Crossing, and from near Halls Creek. Shortest access from Derby lo the north-western part of the Sheet area is by way of Mount House. Within the area graded station tracks radiate to stockyards and bores. All roads in the region are impassable at times during the wet season. The homesteads in the area are served by fortnightly air services from Derby and Wyndham, which have frequent connexions Light aircraft are available for charter in Climate and ~e~et~ti~n The climate is semi-arid and monsoonal, with a short wet s dry winter. The average annual rainfall varies between 18 maxima range from about 8 The natural vegetation consis rock. Large eucalypts an abori~n~s. 12636/61-2 scales of 1 : 250,000 and 1 : 63,360, and a topographic map at 1 : 250,000 scale produced by the Royal Australian Survey Corps are also available. -

Gradual Miocene to Pleistocene Uplift of the Central American Isthmus: Evidence from Tropical American Tonnoidean Gastropods Alan G
J. Paleont., 75(3), 2001, pp. 706±720 Copyright q 2001, The Paleontological Society 0022-3360/01/0075-706$03.00 GRADUAL MIOCENE TO PLEISTOCENE UPLIFT OF THE CENTRAL AMERICAN ISTHMUS: EVIDENCE FROM TROPICAL AMERICAN TONNOIDEAN GASTROPODS ALAN G. BEU Institute of Geological and Nuclear Sciences, P O Box 30368, Lower Hutt, New Zealand, ,[email protected]. ABSTRACTÐTonnoidean gastropods have planktotrophic larval lives of up to a year and are widely dispersed in ocean currents; the larvae maintain genetic exchange between adult populations. They therefore are expected to respond rapidly to new geographic barriers by either extinction or speciation. Fossil tonnoideans on the opposite coast of the Americas from their present-day range demonstrate that larval transport still was possible through Central America at the time of deposition of the fossils. Early Miocene occurrences of Cypraecassis tenuis (now eastern Paci®c) in the Caribbean probably indicate that constriction of the Central American seaway had commenced by Middle Miocene time. Pliocene larval transport through the seaway is demonstrated by Bursa rugosa (now eastern Paci®c) in Caribbean Miocene-latest Pliocene/Early Pleistocene rocks; Crossata ventricosa (eastern Paci®c) in late Pliocene rocks of Atlantic Panama; Distorsio clathrata (western Atlantic) in middle Pliocene rocks of Ecuador; Cymatium wiegmanni (eastern Paci®c) in middle Pliocene rocks of Atlantic Costa Rica; Sconsia sublaevigata (western Atlantic) in Pliocene rocks of Darien, Paci®c Panama; and Distorsio constricta (eastern Paci®c) in latest Pliocene-Early Pleistocene rocks of Atlantic Costa Rica. Continued Early or middle Pleistocene connections are demonstrated by Cymatium cingulatum (now Atlantic) in the Armuelles Formation of Paci®c Panama. -

Historic Initiatives for the Kimberley Saving Australia’S Threatened Wildlife
special edition wildlife matters Autumn 2012 Historic initiatives for the Kimberley Saving Australia’s threatened wildlife Welcome to our special Kimberley edition of Wildlife Matters. The focus of this The AWC mission edition is on two groundbreaking new initiatives in the Kimberley: the Yulmbu project The mission of Australian Wildlife and our historic partnership with the Western Australian Government to protect the Conservancy (AWC) is the effective full length of the Artesian Range. conservation of all Australian animal These two initiatives represent exciting developments for AWC and the wildlife of species and the habitats in which they live. the Kimberley. Most importantly, however, we believe they establish new models To achieve this mission, our actions are with potentially far-reaching consequences for the way on-ground conservation is focused on: delivered, particularly in remote areas of northern and central Australia. • Establishing a network of sanctuaries which protect threatened wildlife and The Yulmbu project was initiated by the indigenous community at Yulmbu, who ecosystems: AWC now manages we have been working with for several years. As part of the project, Yulmbu are 23 sanctuaries covering over 3 million subleasing their land (Tableland Station) to AWC for more than 45 years. It will be hectares (7.4 million acres). managed in accordance with an agreed strategy which encompasses specific targets • Implementing practical, on-ground for feral animal control and fire management. The benefits to Yulmbu are significant: conservation programs to protect they receive annual income (the sublease payments), training and employment in the wildlife at our sanctuaries: these the delivery of land management programs, infrastructure improvements and a programs include feral animal control, modest, sustainable cattle operation. -
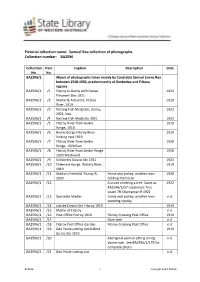
Collection Name: Samuel Rea Collection of Photographs Collection Number: BA2596
Pictorial collection name: Samuel Rea collection of photographs Collection number: BA2596 Collection Item Caption Description Date No. No. BA2596/1 Album of photographs taken mainly by Constable Samuel James Rea between 1918-1932, predominantly of Kimberley and Pilbara regions BA2596/1 /1 Fitzroy to Derby with Native 1921 Prisoners Dec 1921 BA2596/1 /2 Martin & A B Scott, Fitzroy 1919 River, 1919 BA2596/1 /3 Netting Fish Meda Stn, Derby, 1921 1921, Nov BA2596/1 /4 Netting Fish Meda Stn 1921 1921 BA2596/1 /5 Fitzroy River from Geikie 1919 Range, 1919 BA2596/1 /6 Geikie Gorge Fitzroy River 1919 looking east 1919 BA2596/1 /7 Fitzroy River from Geikie 1920 Range, 1920 East BA2596/1 /8 Fitzroy River from Geikie Range 1920 1920 Westward BA2596/1 /9 Kimberley Downs Stn 1921 1921 BA2596/1 /10 Telemere Gorge. Watery River, 1919 1919 BA2596/1 /11 Martin's Emerald. Fitzroy R, Horse and jockey, another man 1920 1920 holding the horse BA2596/1 /12 A snake climbing a tree. Same as 1922 BA2596/1/57 captioned: Tree snake 7ft Champman R 1922 BA2596/1 /13 Scarsdale Martin Horse and jockey, another man n.d. standing nearby BA2596/1 /14 Jubilee Downs Stn Fitzroy 1919 1919 BA2596/1 /15 Martin at Fitzroy n.d. BA2596/1 /16 Post Office Fitzroy 1919 Fitzroy Crossing Post Office 1919 BA2596/1 /17 Gum tree n.d. BA2596/1 /18 Fitzroy Post Office Garden Fitzroy Crossing Post Office n.d. BA2596/1 /19 Geo Poole cutting out bullock 1919 Go Go Stn 1919 BA2596/1 /20 Aboriginal woman sitting on log n.d. -

Catchment Function Analysis Using Google Earth Mapping1
Catchment Function Analysis Wiley Australasian Agribusiness Perspectives 2017, Volume 20, Paper 1 ISSN: 1442-6951 -------------------------------------------------------------------------------------------------------------------------- Catchment Function Analysis using Google Earth Mapping1 Tim Wiley Previously Rangelands NRM, Marble Bar, WA, now Tierra Australia Pty Ltd, Broome, WA. Contact details: [email protected]. -------------------------------------------------------------------------------------------------------------------------- Abstract This paper presents a Catchment Function Analysis utilising Natural Sequence Farming and Ecosystems Management Understanding, as conceptual frameworks for system functioning, together with Google Earth, to indicate critical features of a catchment and the resulting public and private policy implications for catchment management. Key words: catchment function analysis; catchment management; DeGrey; Google mapping. Introduction The importance of understanding the main components of landscapes, and the interrelationships among them, is rising with the threat of significant impacts of climate change and the persistent pressure to lift agricultural productivity while preserving or enhancing ecosystems. Adroit observation of subsystem interactions and the use of new knowledge to scale them up to relevant systems levels will be central to progress in this domain. This paper reports such an approach. The purpose is to explain a novel process for analysing comprehensively a catchment (the -

Rangelands, Western Australia
Biodiversity Summary for NRM Regions Species List What is the summary for and where does it come from? This list has been produced by the Department of Sustainability, Environment, Water, Population and Communities (SEWPC) for the Natural Resource Management Spatial Information System. The list was produced using the AustralianAustralian Natural Natural Heritage Heritage Assessment Assessment Tool Tool (ANHAT), which analyses data from a range of plant and animal surveys and collections from across Australia to automatically generate a report for each NRM region. Data sources (Appendix 2) include national and state herbaria, museums, state governments, CSIRO, Birds Australia and a range of surveys conducted by or for DEWHA. For each family of plant and animal covered by ANHAT (Appendix 1), this document gives the number of species in the country and how many of them are found in the region. It also identifies species listed as Vulnerable, Critically Endangered, Endangered or Conservation Dependent under the EPBC Act. A biodiversity summary for this region is also available. For more information please see: www.environment.gov.au/heritage/anhat/index.html Limitations • ANHAT currently contains information on the distribution of over 30,000 Australian taxa. This includes all mammals, birds, reptiles, frogs and fish, 137 families of vascular plants (over 15,000 species) and a range of invertebrate groups. Groups notnot yet yet covered covered in inANHAT ANHAT are notnot included included in in the the list. list. • The data used come from authoritative sources, but they are not perfect. All species names have been confirmed as valid species names, but it is not possible to confirm all species locations. -

Biodiversity Summary: Rangelands, Western Australia
Biodiversity Summary for NRM Regions Guide to Users Background What is the summary for and where does it come from? This summary has been produced by the Department of Sustainability, Environment, Water, Population and Communities (SEWPC) for the Natural Resource Management Spatial Information System. It highlights important elements of the biodiversity of the region in two ways: • Listing species which may be significant for management because they are found only in the region, mainly in the region, or they have a conservation status such as endangered or vulnerable. • Comparing the region to other parts of Australia in terms of the composition and distribution of its species, to suggest components of its biodiversity which may be nationally significant. The summary was produced using the Australian Natural Natural Heritage Heritage Assessment Assessment Tool Tool (ANHAT), which analyses data from a range of plant and animal surveys and collections from across Australia to automatically generate a report for each NRM region. Data sources (Appendix 2) include national and state herbaria, museums, state governments, CSIRO, Birds Australia and a range of surveys conducted by or for DEWHA. Limitations • ANHAT currently contains information on the distribution of over 30,000 Australian taxa. This includes all mammals, birds, reptiles, frogs and fish, 137 families of vascular plants (over 15,000 species) and a range of invertebrate groups. The list of families covered in ANHAT is shown in Appendix 1. Groups notnot yet yet covered covered in inANHAT ANHAT are are not not included included in the in the summary. • The data used for this summary come from authoritative sources, but they are not perfect. -

Karst Wetlands Biodiversity Ani) Continuity Through Major Climatic Change: an Example from Arid Tropical Western Australia
KARST WETLANDS BIODIVERSITY ANI) CONTINUITY THROUGH MAJOR CLIMATIC CHANGE: AN EXAMPLE FROM ARID TROPICAL WESTERN AUSTRALIA W.F.Humphreys krrestrial InvertebrateZoology, Museum o.f l,{atural Science, WesternAustralian Museum, Francis Street,Perth, WA6000, Australia; E-mail: [email protected]. gov. au Abstract Subterraneankarst wetlands,recently included as a wetland type under the Ramsarclassification, are introducedhere to the wetlandsbiodiversity literature.The generalcharacteristics of karst land- scapesexplain the connectionswith non-karsticwetlands, the often diffuse limits to karst wetlands and their vulnerability to exploitation and pollution. Less than l0oA of the Earth shows distinctive karst landforms but on a global basiskarst wetlandssupport the greaterpart of subterraneanbiodi- versity.This introduction is amplified by a detailed account of a single karst wetland Cape Range peninsula,Western Australia. Cape Range,the only orogenicTertiary limestone in Australia, occurs on a continent relatively devoid of karst. It lies in the arid tropics where surface water flows are mostly episodic and fre- quently saline. The Cape Range karst wetlands contain both freshwater and anchialine wetlands whose natural accessis through a few karst windows and caves.The anchialine system shows marked stratification of their physico-chemicaland biological profiles. The lack of surfacewaters and the particular geographical context means that much of the groundwater fauna represents ancient geographical or phyletic relicts. A general case is made supporting the persistenceof groundwaterfauna in karstic systemsthrough geologicaleras. Introduction The 6th Conferenceof ContractingParties to the RamsarConventionl (Brisbane 1996) voted to include subterraneankarst wetlandsas a wetland type under the Ramsarclassification system. This inclusionhas far reachingimplications, not only for karst regions,but also for a wide rangeof wetlands,and for groundwatercon- servationgenerally.