ERMA200599 Application Appendix 3.Pdf
Total Page:16
File Type:pdf, Size:1020Kb
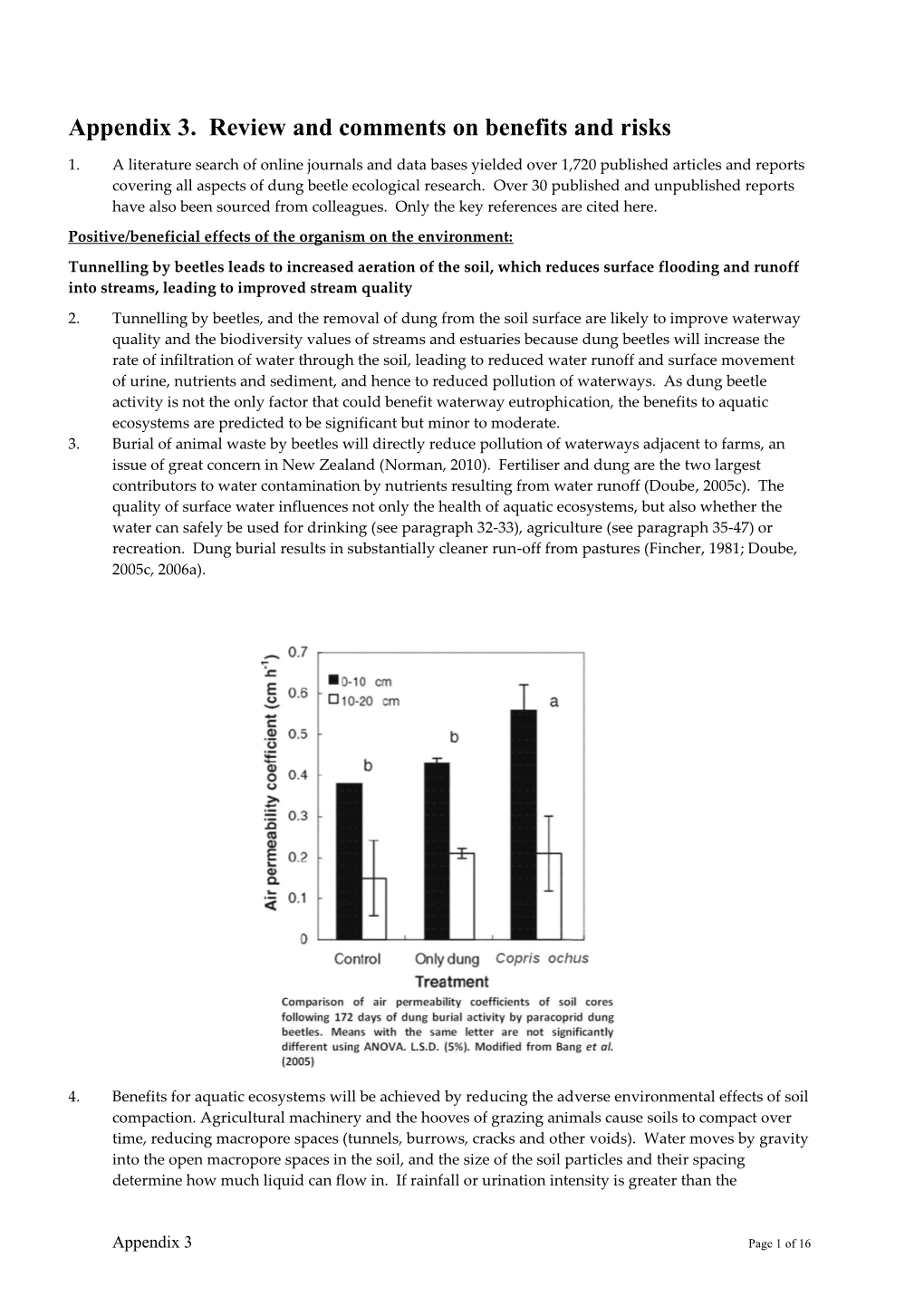
Load more
Recommended publications
-

Phylogenetic Analysis of Geotrupidae (Coleoptera, Scarabaeoidea) Based on Larvae
Systematic Entomology (2004) 29, 509–523 Phylogenetic analysis of Geotrupidae (Coleoptera, Scarabaeoidea) based on larvae JOSE´ R. VERDU´ 1 , EDUARDO GALANTE1 , JEAN-PIERRE LUMARET2 andFRANCISCO J. CABRERO-SAN˜ UDO3 1Centro Iberoamericano de la Biodiversidad (CIBIO), Universidad de Alicante, Spain; 2CEFE, UMR 5175, De´ partement Ecologie des Arthropodes, Universite´ Paul Vale´ ry, Montpellier, France; and 3Departamento Biodiversidad y Biologı´ a Evolutiva, Museo Nacional de Ciencias Naturales (CSIC), Madrid, Spain Abstract. Thirty-eight characters derived from the larvae of Geotrupidae (Scarabaeoidea, Coleoptera) were analysed using parsimony and Bayesian infer- ence. Trees were rooted with two Trogidae species and one species of Pleocomidae as outgroups. The monophyly of Geotrupidae (including Bolboceratinae) is supported by four autapomorphies: abdominal segments 3–7 with two dorsal annulets, chaetoparia and acanthoparia of the epipharynx not prominent, glossa and hypopharynx fused and without sclerome, trochanter and femur without fossorial setae. Bolboceratinae showed notable differences with Pleocomidae, being more related to Geotrupinae than to other groups. Odonteus species (Bolboceratinae s.str.) appear to constitute the closest sister group to Geotrupi- nae. Polyphyly of Bolboceratinae is implied by the following apomorphic char- acters observed in the ‘Odonteus lineage’: anterior and posterior epitormae of epipharynx developed, tormae of epipharynx fused, oncyli of hypopharynx devel- oped, tarsal claws reduced or absent, plectrum and pars stridens of legs well developed and apex of antennal segment 2 with a unique sensorium. A ‘Bolbelas- mus lineage’ is supported by the autapomorphic presence of various sensoria on the apex of the antennal segment, and the subtriangular labrum (except Eucanthus). This group constituted by Bolbelasmus, Bolbocerosoma and Eucanthus is the first evidence for a close relationship among genera, but more characters should be analysed to test the support for the clade. -

Seasonality of Coprophagous Beetles in the Kaiserstuhl Area Near Freiburg (SW-Germany) Including the Winter Months
1994, 15 (5), 607-631 Seasonality of coprophagous beetles in the Kaiserstuhl area near Freiburg (SW-Germany) including the winter months Thomas Waßmer Institute of Landscape Ecology and Landuse Planning, Faculty of Forestry, Albert-Ludwigs-University of Freiburg D-79085 Freiburg. Current address: Lehrstuhl Markl, Fakultät fur Biologie, Universität Konstanz, Postfach 5560 M618, D-78434 Konstanz, Germany. Abstract In an all-year-round survey of coprophagous beetles in a pasture in the Kaiserstuhl area (SW-Germany), dung pats were gathered twice a month (on the 1st and the 15th). 40,298 beetles belonging to 38 species of Scarabaeoidea and 14 of Hydrophilidac, were detected. The small site on the Kaiserstuhl has the highest species diversity in Scarabaeoidea species of all local dung beetle coenoses described in Europe so far. A comparison of monthly dominance structures of the species community between the Kaiserstuhl and warm temperate pastures in the south of Spain yields similarities on both the subgenus and species levels. Analyses of the phenological distributions of 34 abundant dung beetle species show a high degree of specialisation among all taxonomic and phenologic groups, most prominently among the Aphodius. There are even species which occur predominantly during the winter months, which might be due to low interspecific competition in these months and favourable synchronisation of mating and reproduction. Cluster analysis confirms eight phenological groups and proves the very structuring nature of the niche dimension season. Beside the qualitative separation of species, there was remarkable segregation in the biomass of species in the different months. "Dwellers" (endocoprid species) of the subfamily Aphodiinae showed the most pronounced separation, followed by small "Tunnelers" (paracoprid species) belonging to the Onthophagus big "Tunnelers" of the Copris and Geotrupes and small Hydrophilidae "Dwellers" of the genus Cercyon. -

Reproductive Behaviour and Development of the Dung Beetle Typhaeus Typhoeus (Coleoptera, Geotrupidae)
REPRODUCTIVE BEHAVIOUR AND DEVELOPMENT OF THE DUNG BEETLE TYPHAEUS TYPHOEUS (COLEOPTERA, GEOTRUPIDAE) by LIJBERTBRUSSAARD Dept. of Animal Ecology and Dept. of Soil Science & Geology, Agricultural University, Wageningen, The Netherlands . ABSTRACT This paper is part of a study of the contribution of dung beetles to soil formation in sandy soils. Typhaeus typhoeus (Linnaeus) has been selected because it makes deep burrows and is locally abundant. The beetles are active from autumn until spring, reproduction takes place from February to April. Sex pheromones probably influence pair formation. The sexes co operate in excavating a burrow (up to 0.7 m below surface) and in provisioning the burrow with dung as food for the larvae. Co-operation is reset by scraping each other across the thorax or elytra. Dung sausages, appr. 12.5 cm long and 15 mm in diameter, are manufac tured above each other. Development is rapid at 13—17°C. The life cycle is accelerated by a cold period in the third larval stage. These requirements are met by soil temperatures up to 15° C in summer and down to 5 °C in winter. The life cycle lasts two years, but longer under certain conditions. Newly hatched beetles make their way to the surface through the soil, but do not follow the old shaft. Adults reproduce only once. Differential rate of com pletion of the life cycle and occasional flying probably reduce the risk of local extinction. The study is thought to be relevant for behavioural ecology and soil science. CONTENTS tion of how much dung beetles contribute to Introduction 203 soil formation today. -

The Role of Semi–Natural Grasslands and Livestock in Sustaining Dung Beetle Communities (Coleoptera, Scarabaeoidea) in Sub–Mediterranean Areas of Slovenia
Animal Biodiversity and Conservation 41.2 (2018) 321 The role of semi–natural grasslands and livestock in sustaining dung beetle communities (Coleoptera, Scarabaeoidea) in sub–Mediterranean areas of Slovenia J. Jugovic, N. Koprivnikar, T. Koren Jugovic, J., Koprivnikar, N., Koren, T., 2018. The role of semi–natural grasslands and livestock in sustaining dung beetle communities (Coleoptera, Scarabaeoidea) in sub–Mediterranean areas of Slovenia. Animal Biodiversity and Conservation, 41.2: 321–332, Doi: https://doi.org/10.32800/abc.2018.41.0321 Abstract The role of semi–natural grasslands and livestock in sustaining dung beetle communities (Coleoptera, Scara- baeoidea) in sub–Mediterranean areas of Slovenia. We studied the richness and structure of the coprophagous Scarabaeoidea community in two pastures (Hrastovlje and Zazid) in sub–Mediterranean Slovenia. In each pasture, we examined three habitat patches characterised by different levels of grazing (S1, the active part of the pasture; S2, the overgrown part of the pasture, mainly spiny shrubs; S3, a meadow with some overgrown patches of shrubs outside the fenced pasture). The main results were as follows: (1) 29 species were sampled, corresponding to about three quarters of the species presumably present at the two study sites; (2) species richness and abundance in Zazid are were similar in all three patches; (3) the species richness and abundance in Hrastovlje (in total, and separately for dwellers and tunnelers) were highest in S2. In Hrastovlje, dwellers were most abundant in S1. As the two different habitat patches were shown to positively influence the dung beetle community, we recommend maintaining a traditionally–managed mosaic landscape. -

Experimental Addition of Onthophagus Binodis and Geotrupes Spiniger to Pasture Ecosystems in New Zealand and an Investigation Of
Experimental addition of Onthophagus binodis and Geotrupes spiniger to pasture ecosystems in New Zealand and an investigation of affected biotic and abiotic soil factors. Rebecca L. Sewell A thesis submitted in fulfilment of the requirements of the degree of Master of Science in Zoology at the University of Otago, Dunedin New Zealand 2018 ii Abstract New Zealand has a serious environmental pollution problem caused by intensive farming, and a lack of co-evolved dung beetle species to clean up the waste produced by these introduced herbivores. Our endemic dung beetles evolved in isolation from large herbivores such as cattle, sheep and horses and consequently don’t use the dung of these species. New exotic species of dung beetle have been introduced into New Zealand with the purpose of establishing populations that will combat this agricultural pollution. All of the species approved for introduction share the trait of being paracoprid or tunnelling. They dig tunnels below the dung pat and construct brood chambers from dung brought down from the surface. Significant benefits from this behaviour have been documented in other countries where these beetles are already established, however there is a paucity of studies documenting these effects in New Zealand soils. The value of this research lies in the investigation of exotic dung beetles in novel soil types. Here we aimed to demonstrate in Otago soils the benefits to soil physical structure, nutrient profiles, and invertebrate species (particularly earthworms) which have been found in other environments. A field study was conducted from February to June 2015 at two Otago sites – one a lowland intensive farming system (Mosgiel) and the other an upland less modified system (near Middlemarch) with some native tussock grass. -

Appendix 4. Application References
Appendix 4. Application References AgResearch (2010). Managing parasite resistance to anthelmintic drugs. Science Review. http://www.agresearch.co.nz/publications/sciencereview/parasite-resistance.pdf. Retrieved 25 August, 2010. Bang HS, Lee J-H, Kwon OS, Na YE, Jang YS, Kim WH. (2005). Effects of paracoprid dung beetles (Coleoptera: Scarabaeidae) on the growth of pasture herbage and on the underlying soil. Applied Soil Ecology 29: 165–171. Bishop DM. (1998). Parasitic Hyrnenoptera reared from dung-breeding Diptera in New Zealand. New Zealand Entomologist 21: 99- 106. Bishop DM, Heath ACG. (1995). Aphaereta aotea, a hymenopteran parasitoid with the potential to act as a biological control agent agzinst flystrike. New Zealand Journal of Zoology 22: 187 (Abstract). Bishop DM, Heath ACG, Haack NA. (1996). Distribution, prevalence and host associations of hymenoptera parasitic on Calliphoridae occurring in flystrike in New Zealand. Medical and Veterinary Entomology 10: 365-370. Bornemissza GF. (1960). Could dung eating insects improve our pastures? Journal of the Australian Institute of Agricultural Science 26: 54-56. Bornemissza GF. (1970). Insectary studies on the control of dung breeding flies by the activity of the dung beetle, Onthophagus gazella F. (Coleoptera: Scarabaeinae). Journal of the Australian Entomological Society 9: 31-41. Bornemissza GF. (1976). The Australian dung beetle project 1965-1975. Australian Meat Research Committee Review 30: 32pp. Bornemissza GF, Williams CH. (1970). An effect of dung beetle activity on plant yield. Pedobiologia 10: 1-7. Blume RR, Matter JJ, Eschle JL. (1973). Onthophagus gazella: effect on survival of horn flies in the laboratory. Environmental Entomology 2: 811-813. Bryan RP. (1973). -

ERMA New Zealand Evaluation and Review Report
ERMA New Zealand Evaluation and Review Report Application to release 11 species of dung beetle to overcome the many adverse effects caused by animal dung in New Zealand pastures Application Code: ERMA200599 Prepared for the Environmental Risk Management Authority 1. Executive Summary A consortium of farmers, researchers and other interested parties has applied to the Environmental Risk Management Authority (the Authority) to release eleven species of dung beetle. This evaluation and review report is written by the Agency, a body which supports the Authority. It provides an overview of the beneficial and adverse effects of releasing these eleven species. The Agency has found that there are multiple non-negligible benefits to releasing these species. There are environmental and human health benefits from enhanced water and soil quality and economic benefits from an increase in farm productivity by the actions of the beetles. Given the obligate nature of true dung beetles in regard to what they feed on, one of the beetles in the application has been suggested to be declined. Of the proposed dung beetles, Geotrupes spiniger is in a Family that are known to utilise other food sources besides herbivore dung and thus poses a non-negligible risk to the native Scarab beetles. Of the remaining ten species no adverse effects were found to be significant given that the recommended species will be restricted to pastureland and are reliant on large herbivore dung. The Agency is therefore recommending approval of all the dung beetles in this application to import for release except Geotrupes spiniger. ERMA New Zealand Evaluation & Review Report: ERMA200599 Page | 2 2. -

A Review of the Status of the Beetles of Great Britain
Natural England Commissioned Report NECR224 A review of the status of the beetles of Great Britain The stag beetles, dor beetles, dung beetles, chafers and their allies - Lucanidae, Geotrupidae, Trogidae and Scarabaeidae Species Status No.31 First published 31 October 2016 www.gov.uk/natural-england Foreword Natural England commission a range of reports from external contractors to provide evidence and advice to assist us in delivering our duties. The views in this report are those of the authors and do not necessarily represent those of Natural England. Background Decisions about the priority to be attached to the conservation of species should be based upon objective assessments of the degree of threat to species. The internationally-recognised approach to undertaking this is by assigning species to one of the IUCN threat categories using the IUCN guidelines. This report was commissioned to update the national threat status of beetles within the Lucanidae, Geotrupidae, Trogidae and Scarabaeidae. It covers all species in these groups, identifying those that are rare and/or under threat as well as non-threatened and non- native species. Reviews for other invertebrate groups will follow. Natural England Project Manager – Jon Webb, [email protected] Contractor – Steve Lane [email protected] Authors – Steve A. Lane & Darren J. Mann Keywords – Scarabaeidae, Lucanidae, Geotrupidae, Trogidae, chafers, dung beetles, stag beetles, dor beetles, rhinoceros beetle, invertebrates, red list, IUCN, status reviews Further information This report can be downloaded from the Natural England Access to Evidence Catalogue: http://publications.naturalengland.org.uk/. For information on Natural England publications contact the Natural England Enquiry Service on 0300 060 3900 or e-mail [email protected]. -

Dung Feeding in Hydrophilid, Geotrupid and Scarabaeid Beetles: Examples of Parallel Evolution
Eur. J. Entomol. 101: 365–372, 2004 ISSN 1210-5759 Dung feeding in hydrophilid, geotrupid and scarabaeid beetles: Examples of parallel evolution PETER HOLTER Department of Terrestrial Ecology, Institute of Biology, University of Copenhagen, Universitetsparken 15, DK-2100 Copenhagen Ø, Denmark; e-mail: [email protected] Key words. Dung beetles, Hydrophilidae, Geotrupidae, Scarabaeidae, feeding habits, particle feeding, mouthparts, evolution Abstract. The maximum size of ingested ball-shaped particles was determined in three species of adult dung feeding beetle: Anoplo- trupes (Geotrupes) stercorosus and Geotrupes spiniger (Geotrupidae, Geotrupinae) and Sphaeridium lunatum (Hydrophilidae, Sphaeridiinae). Maximum diameters were 40–65 µm, 60–75 µm and 16–19 µm in A. stercorosus, G. spiniger and S. lunatum, respectively, and it was concluded that these beetles feed in the same way as found in previous studies on coprophagous scarabaeids (Scarabaeinae and Aphodiinae). Coarse particles, mainly indigestible plant fragments, are rejected by an unknown filtering mecha- nism, and only very small particles are actually ingested. The two geotrupids, however, tolerate somewhat larger particles than do scarabaeines of similar size. This may reflect a lower degree of specialisation towards dung feeding in the geotrupids than in the scarabaeines. In several ways, the mouthparts of the coprophagous Scarabaeidae, Geotrupidae and Hydrophilidae show essentially the same morphological modifications that must be adaptations for dung feeding. For the hydrophilid (Sphaeridium), such modifica- tions are described for the first time. They include asymmetric mandibular molars (right convex, left concave), fitting exactly into each other, with highly specialised surfaces that may concentrate the food prior to ingestion by squeezing fluid out of it. -

Type of the Paper (Article
insects Article Diversity Patterns of Dung Beetles along a Mediterranean Elevational Gradient Cristina Mantoni 1, Noelline Tsafack 2 , Ettore Palusci 1, Stefano Di Pietro 1 and Simone Fattorini 1,* 1 Department of Life, Health and Environmental Sciences, University of L’Aquila, Via Vetoio, 67100 L’Aquila, Italy; [email protected] (C.M.); [email protected] (E.P.); [email protected] (S.D.P.) 2 cE3c-Centre for Ecology, Evolution and Environmental Changes, Azorean Biodiversity Group, Faculdade de Ciências e Engenharia do Ambiente, Universidade dos Açores, rua Capitão João D’Avila, 9700-042 Angra do Heroísmo, Portugal; [email protected] * Correspondence: [email protected] Simple Summary: Mountains are ideal natural laboratories to study how biodiversity is influenced by environmental characteristics because climate varies rapidly from lowlands to high elevations. Scientists have investigated how the number of species varies with elevation for the most disparate plant and animal groups worldwide. However, species richness is only one aspect of biodiversity that does not consider species abundances. The so-called Hill numbers are a unified family of mathematical indices that express biodiversity in terms of both richness and abundance. We used Hill numbers to investigate how dung beetle diversity varies along an elevational gradient in a Mediterranean mountain. We found that scarabaeids were the most abundant dung beetle group. These insects construct subterranean nests protecting their offspring from desiccation during warm and dry summer climatic conditions. Additionally, in accordance with their preference for open Citation: Mantoni, C.; Tsafack, N.; habitats, we found that dung beetles are more abundant and diversified in grasslands than in Palusci, E.; Di Pietro, S.; Fattorini, S. -

Ancient Origin of Endemic Iberian Earth-Boring Dung Beetles (Geotrupidae) ⇑ Regina L
Molecular Phylogenetics and Evolution 59 (2011) 578–586 Contents lists available at ScienceDirect Molecular Phylogenetics and Evolution journal homepage: www.elsevier.com/locate/ympev Ancient origin of endemic Iberian earth-boring dung beetles (Geotrupidae) ⇑ Regina L. Cunha a,b, José R. Verdú c, Jorge M. Lobo a, , Rafael Zardoya a a Departamento de Biodiversidad y Biología Evolutiva, Museo Nacional de Ciencias Naturales-CSIC, José Gutiérrez Abascal, 2, 28006 Madrid, Spain b CCMAR, Campus de Gambelas, Universidade do Algarve, 8005-139 Faro, Portugal c Centro Iberoamericano de la Biodiversidad (CIBIO), Universidad de Alicante, San Vicente del Raspeig, E-03080 Alicante, Spain article info abstract Article history: The earth-boring dung beetles belong to the family Geotrupidae that includes more than 350 species clas- Received 9 July 2010 sified into three subfamilies Geotrupinae, Lethrinae, and Taurocerastinae, mainly distributed across tem- Revised 25 February 2011 perate regions. Phylogenetic relationships within the family are based exclusively on morphology and Accepted 29 March 2011 remain controversial. In the Iberian Peninsula there are 33 species, 20 of them endemic, which suggests Available online 5 April 2011 that these lineages might have experienced a radiation event. The evolution of morphological adaptations to the Iberian semi-arid environments such as the loss of wings (apterism) or the ability to exploit alter- Keywords: native food resources is thought to have promoted diversification. Here, we present a phylogenetic anal- Molecular phylogeny ysis of 31 species of Geotrupidae, 17 endemic to the Iberian Peninsula, and the remaining from Geotrupidae Paleoendemism southeastern Europe, Morocco, and Austral South America based on partial mitochondrial and nuclear Apterism gene sequence data. -

Effects of Different Pastures Management on Dung Beetle
UNIVERSITÀ DEGLI STUDI DI URBINO “CARLO BO” DIPARTIMENTO DI SCIENZE PURE E APPLICATE CORSO DI DOTTORATO DI RICERCA IN “SCIENZE AMBIENTALI” CICLO XXVIII Tesi in cotutela: Università degli Studi di Urbino “Carlo Bo” e “Universidad de Alicante” Effects of different pastures management on dung beetle communities in a sub-mountainous landscape of central Italy: a multicomponent biodiversity and ecological process analysis Settore Scientifico Disciplinare: BIO/05 DOTTORANDO: Dott. Mattia Tonelli RELATORI: Prof. Mario Zunino Prof. José Ramón Verdú Faraco Dipartimento di Scienze Pure e Centro Iberoamericano de la Applicate Biodiversidad Università degli Studi di Urbino “Carlo CIBIO (Universidad de Alicante) Bo” ANNO ACCADEMICO 2015/2016 M. Tonelli Effects of different grazing management on dung beetle biodiversity and ecological process M. Tonelli Effects of different grazing management on dung beetle biodiversity and ecological process A te vengo, balena che tutto distruggi ma non vinci: fino all’ultimo lotto con te; dal cuore dell’inferno ti trafiggo; in nome dell’odio, vomito a te l’ultimo mio respiro (Herman Melville) …nulla si può tentare se non stabilire l’inizio e la direzione di una strada infinitamente lunga. La pretesa di qualsiasi completezza sistematica e definitiva sarebbe, se non altro, un’illusione. Qui il singolo ricercatore può ottenere la perfezione solo nel senso soggettivo che egli comunichi tutto ciò che è riuscito a vedere. (Georg Simmel) M. Tonelli Effects of different grazing management on dung beetle biodiversity and ecological process M. Tonelli Effects of different grazing management on dung beetle biodiversity and ecological process Al coraggio di un pettirosso da combattimento M. Tonelli Effects of different grazing management on dung beetle biodiversity and ecological process M.