Conjoined Twinning & Biological Individuation
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

The Human Chimera: Legal Problems Arising from Individuals with Multiple Types of Dna
Seton Hall University eRepository @ Seton Hall Law School Student Scholarship Seton Hall Law 5-2-2014 The umH an Chimera: Legal Problems Arising From Individuals with Multiple Types of DNA Robert Russell Granzen Follow this and additional works at: https://scholarship.shu.edu/student_scholarship Recommended Citation Granzen, Robert Russell, "The umH an Chimera: Legal Problems Arising From Individuals with Multiple Types of DNA" (2014). Law School Student Scholarship. 485. https://scholarship.shu.edu/student_scholarship/485 THE HUMAN CHIMERA: LEGAL PROBLEMS ARISING FROM INDIVIDUALS WITH MULTIPLE TYPES OF DNA Robert Granzen INTRODUCTION Science continually changes, and with it our understanding of the human body. While some scientific developments are limited in scope, others have widespread effects. Scientists have just recently begun understanding the range of effects chimerism in humans can have. Chimerism, originally associated with hermaphrodites having both male and female sexual organs, is much more common than originally thought. As chimerism becomes more common, so do individuals with separate and distinct deoxyribonucleic acid (DNA) strands in their bodies. Most individuals are unaware of their chimeric genetic code and most will likely never know. Because of the inherent difficulty of testing for chimerism, many problems are presented to legal system. Part I of this article will begin with the history of human chimeras. The section will then describe the ways in which chimeras are formed. The section will discuss the most common form of chimerism in humans--fetal cell microchimerism (FMC). FMC occurs when cells are transferred from baby to mother or mother to baby via the umbilical cord.1 Studies have shown that mothers may keep cells from their children for years after giving birth.2 Moreover, cells can be exchanged between twins while inside the uterus.3 Secondly, the section will describe the process of embryo fusion, which can cause tetragametic chimerism. -

Representations of Conjoined Twins
University of Wollongong Research Online University of Wollongong Thesis Collection 1954-2016 University of Wollongong Thesis Collections 2015 Representations of conjoined twins Claire Marita Fletcher University of Wollongong Follow this and additional works at: https://ro.uow.edu.au/theses University of Wollongong Copyright Warning You may print or download ONE copy of this document for the purpose of your own research or study. The University does not authorise you to copy, communicate or otherwise make available electronically to any other person any copyright material contained on this site. You are reminded of the following: This work is copyright. Apart from any use permitted under the Copyright Act 1968, no part of this work may be reproduced by any process, nor may any other exclusive right be exercised, without the permission of the author. Copyright owners are entitled to take legal action against persons who infringe their copyright. A reproduction of material that is protected by copyright may be a copyright infringement. A court may impose penalties and award damages in relation to offences and infringements relating to copyright material. Higher penalties may apply, and higher damages may be awarded, for offences and infringements involving the conversion of material into digital or electronic form. Unless otherwise indicated, the views expressed in this thesis are those of the author and do not necessarily represent the views of the University of Wollongong. Recommended Citation Fletcher, Claire Marita, Representations of conjoined twins, Master of Arts in Creative Writing thesis, Faculty of Law, Humanities and the Arts, University of Wollongong, 2015. https://ro.uow.edu.au/theses/ 4691 Research Online is the open access institutional repository for the University of Wollongong. -

Patentability of Human-Animal Chimeras Ryan Hagglund
Santa Clara High Technology Law Journal Volume 25 | Issue 1 Article 4 2008 Patentability of Human-Animal Chimeras Ryan Hagglund Follow this and additional works at: http://digitalcommons.law.scu.edu/chtlj Part of the Law Commons Recommended Citation Ryan Hagglund, Patentability of Human-Animal Chimeras, 25 Santa Clara High Tech. L.J. 51 (2012). Available at: http://digitalcommons.law.scu.edu/chtlj/vol25/iss1/4 This Article is brought to you for free and open access by the Journals at Santa Clara Law Digital Commons. It has been accepted for inclusion in Santa Clara High Technology Law Journal by an authorized administrator of Santa Clara Law Digital Commons. For more information, please contact [email protected]. PATENTABILITY OF HUMAN-ANIMAL CHIMERAS Ryan Hagglundt Abstract The chimera was a mythological creature with a lion 's head, goat's body, and serpent's tail. Because of recent advances in biotechnology, such permutations on species are no longer the stuff of myth and legend The term "chimera " has come to describe a class of genetically engineered creatures composed of some cells from one species, which thus contain genetic material derived entirely from that species, and some cells from another species, containing only genetic material from that species. Scientists have created a goat- sheep chimera or "geep" which exhibits physical characteristicsof both animals. Likewise, scientists have also used the tools of modern molecular biology to create human-animal chimeras containing both human and animal cells. While none of the human-animal chimeras hitherto created have exhibited significant human characteristics,the synthesis of human-animal chimeras raises significant ethical concerns. -

The Role of Rapid Tissue Expansion in Separating Xipho-Omphalopagus Conjoined Twins in Vietnam
Breast/Trunk Case Report The role of rapid tissue expansion in separating xipho-omphalopagus conjoined twins in Vietnam Tran Thiet Son1,2,3, Pham Thi Viet Dung1, Ta Thi Hong Thuy1, Vu Duy Kien4, Nguyen Thanh Liem5 1Department of Plastic and Reconstructive Surgery, Hanoi Medical University Hospital, Hanoi; 2Department of Plastic Reconstructive Aesthetic Surgery, Saint Paul Hospital, Hanoi; 3Department of Plastic and Reconstructive Surgery, Bach Mai Hospital, Hanoi; 4OnCare Medical Technology Company Limited, Hanoi; 5Department of Pediatric Surgery, Vietnam National Children’s Hospital, Hanoi, Vietnam Conjoined twins are rare, and each set of conjoined twins has a unique conjoined anatomy. It Correspondence: Tran Thiet Son is necessary to perform separation to increase the chance of patient survival. Tissue expan- Department of Plastic and Reconstructive Surgery, Hanoi sion is an advanced technique for providing sufficient soft tissue and skin for wound closure. Medical University Hospital, No.1 Ton We report the successful application of rapid tissue expansion in 10-month-old xipho-om- That Tung Street, Hanoi 116001, phalopagus conjoined twins in Vietnam. A tissue expander was placed on the anterior body Vietnam between the sternum and umbilicus with a baseline of 70 mL sterile saline (0.9% NaCl). The Tel: +84-903444244 E-mail: [email protected] first injection into the tissue expander began on the 6th day after expander insertion, and in- jections continued every 2 days with approximately 30–70 mL per injection according to the expansion of the skin. The expander reached 335 mL after six injections and within 10 days. In order to prepare for surgical separation, expansion was completed on the 15th day after insertion. -

Twin-To-Twin Transfusion Syndrome: an Overview Richa Saxena1, Kanav Midha2
REVIEW ARTICLE Twin-to-twin Transfusion Syndrome: An Overview Richa Saxena1, Kanav Midha2 ABSTRACT Twin-to-twin transfusion syndrome (TTTS) is a severe problem that affects 10–15% of monochorionic (MC) multiple pregnancies. Connecting placental vessels on chorionic plate between donor and recipient twin is accountable for inequality of blood flow. There is an indication for the superiority of fetoscopic laser ablation over serial amnioreductions regarding survival and neurological outcome for stages II–IV TTTS. However, the optimal management of stage I is still debated. In this review, we discuss the basics of twin gestation, optimal management, pathophysiology, long-term neurodevelopmental outcome, and future aspects of TTTS. Keywords: Dizygotic, Fetus, Gestation, Monozygotic, Twin–twin transfusion syndrome. World Journal of Anemia (2018): 10.5005/jp-journals-10065-0041 INTRODUCTION 1Jaypee Brothers Medical Publishers, New Delhi, India Development of two or more embryos simultaneously in a pregnant 2Department of Pharmaceutical Sciences, Chitkara University, uterus is termed as “multifetal gestation.” Development of two Chandigarh, India fetuses (whether through monozygotic or dizygotic fertilization) Corresponding Author: Richa Saxena, Jaypee Brothers Medical simultaneously is known as twin gestation; development of three Publishers, New Delhi, India, Phone: +91 9971234834, e-mail: synapse94@ fetuses simultaneously as triplets; four fetuses as quadruplets; five hotmail.com fetuses as quintuplets, and so on. The incidence of twin gestation How to cite this article: Saxena R, Midha K. Twin-to-twin Transfusion is about 1 per 80 live births. The incidence varies among different Syndrome: An Overview. World J Anemia 2018;2(3–4):96–102. countries and ethnic groups, with the incidence being highest Source of support: Nil in African countries, lowest in Japan and intermediate among Conflict of interest: None Caucasians. -

My Attempt to Patent a Human-Animal Chimera
No 27 - avril-mai 2006 My attempt to patent a human-animal chimera In 1997, with the support of the social critic Jeremy Rifkin of the Foundation on Economic Trends in Washington, D.C., Professor Stuart A. Newman undertook to apply for a patent on embryos and full- term creatures containing human along with nonhuman cells – so-called “chimeras” that he had no intention of producing... Stuart A. Newman* In 1997, with the support of the social critic Jeremy Rifkin of the Foundation on Economic Trends in Washington, D.C., I undertook to apply for a patent on embryos and full-term creatures containing human along with nonhuman cells – so-called “chimeras”. The public had only just learned (to widespread surprise) about the cloning of Dolly the sheep and the announcement of the production of versatile and controversial embryonic stem (ES) cells from human embryos was still a year in the future. In those innocent days less than a decade ago the prospect of fabricating creatures that were part-human and part-beast would have appeared to most people like something out of Greek mythology or the pages of a fantasy novel. Indeed, it had been a century since the British writer H.G. Wells had published “The Island of Dr. Moreau”, the story of a visionary scientist whose well-motivated experiments along these lines led to death and destruction. Still, in the last years of the 20th century, such quasi-humans were unknown outside of works of fiction. Nonetheless, as a developmental biologist whose work requires close tracking of the scientific literature, I knew that such things were feasible. -

Parapagus Conjoined Twins: Complicated Anatomy Precludes Separation
Case Report Full text online at http://www.jiaps.com Parapagus conjoined twins: Complicated anatomy precludes separation Arbinder K. Singhal, Gautam S. Agarwal, Shilpa Sharma, Arun K. Gupta*, Devendra K. Gupta Departments of Pediatric Surgery and *Radiodiagnosis, All India Institute of Medical Sciences, New Delhi, India Correspondence: Dr. D. K. Gupta, Department of Pediatric Surgery, All India Institute of Medical Sciences, New Delhi - 110 029, India. E-mail: [email protected] ABSTRACT A parapagus set of male conjoined twins was brought to our institution at 12 h after birth. An extensive sharing of the abdominal viscera (single liver, hindgut), abdominal aorta, pelvis (single rectum and anus), genitalia (one set) and vertebral column was found. The surgical separation was not considered due to medical and ethical issues. KEY WORDS: Parapagus, conjoint twins INTRODUCTION were cyanosed. The right twin was intubated and required ventilation for 24 h; after that, both the babies Conjoined twins occur in one in 50-100,000 live births remained stable. and are now thought to result from the fission of the notochordal anlage; followed by subsequent fusion of Babygram showed two separate vertebral columns with the embryos.[1] Parapagus twins joined anterolaterally scoliosis and separate hemisacra, which joined distally result from two nearly parallel notochords in close [Figure 2]. Echocardiogram, ultrasonogram with doppler proximity. This anomaly represents less than 0.5% of and contrast enhanced computed tomographic (CECT) all reported cases of conjoined twins.[2] We present one scan revealed two structurally normal hearts with liver such surviving set of conjoined twins. wedged in between; two separate thoracic aortas; two separate inferior vena cavae with ipsilateral drainage of MATERIALS AND METHODS hepatic veins; a central large liver with a single gall bladder; single spleen on the left side; and two normal A set of male conjoined twins, born to a third gravida kidneys, one in each flank. -

The First Successful Separation of Conjoined Twins in 1689: Some Additions and Corrections
The First Successful Separation of Conjoined Twins in 1689: Some Additions and Corrections Erwin J. O. Kompanje Department of Neurological Surgery, Erasmus Medical Center Rotterdam, the Netherlands he surgical separation of a pair of conjoined twins in Curiosa sive Ephemeridum Medico-Physicarum Tthe year 1689 by Johannes Fatio was the subject of Germanicarum Academiae Imperialis Leopoldinae a recent article in this journal. The reference used as Naturae Curiosorum (König 1689; Zwinger 1690). publication of the case was Fatio’s book, Der Arzney These two articles must be considered as the two ear- Doctor, Helvetisch-Vernüftige Wehe-Mutter, published liest sources concerning this case. The article of König in 1752, although the case was presented in literature (König, 1689) is illustrated with the same engraved by three earlier sources. Two articles were published plate (Figure 1) as Fatio (Figure 3), with some differ- in the Miscellanea Curiosa sive Ephemeridum Medico- ences. König’s plate can be considered as the first Physicarum Germanicarum Academiae Imperialis engraving of the case, leaving Fatio’s plate as a copy Leopoldinae Naturae Curiosorum in 1689 by Emanuel from this engraving. In 1689 Johannes Fatio provided König and another in 1690 by Theodor Zwinger who described and illustrated the case in detail. Besides the details of the case to Zwinger, which formed the these articles, a Flug Blatt was published on the case basis of Zwinger’s article. The girls depicted on the between 1689 and 1695. Fatio copied the engraved engraving were named Elisabet (Elisabetham) and plate in his book from König’s engraving. These two Catherina (Catharinam). -

Separating Conjoined Twins: Legal Reverberations of Jodie and Mary's Predicament
Loyola of Los Angeles International and Comparative Law Review Volume 24 Number 1 Article 3 1-1-2002 Separating Conjoined Twins: Legal Reverberations of Jodie and Mary's Predicament Jennifer N. Sawday Follow this and additional works at: https://digitalcommons.lmu.edu/ilr Part of the Law Commons Recommended Citation Jennifer N. Sawday, Separating Conjoined Twins: Legal Reverberations of Jodie and Mary's Predicament, 24 Loy. L.A. Int'l & Comp. L. Rev. 65 (2002). Available at: https://digitalcommons.lmu.edu/ilr/vol24/iss1/3 This Notes and Comments is brought to you for free and open access by the Law Reviews at Digital Commons @ Loyola Marymount University and Loyola Law School. It has been accepted for inclusion in Loyola of Los Angeles International and Comparative Law Review by an authorized administrator of Digital Commons@Loyola Marymount University and Loyola Law School. For more information, please contact [email protected]. NOTES AND COMMENTS SEPARATING CONJOINED TWINS: LEGAL REVERBERATIONS OF JODIE AND MARY'S PREDICAMENT On December 10, 2000, a coroner recorded a special verdict for the death of a baby girl in England. 1 The coroner faced an unusual situation - a death from surgery when it was known beforehand the patient was going to die.2 The coroner admitted that all surgeries carry a risk of mortality, but in this case the mortality rate from the surgery was known beforehand to be one hundred percent.3 The special verdict read: "Mary died following surgery separating her from her conjoined twin, which surgery was permitted by an order of the [British] High Court, confirmed by the Court of Appeal."'4 I. -

Chimera Proposal
Frequently Asked Questions on Chimera Proposal 1) What are the potential benefits of this research? Research using animal models containing human cells is certainly not new. For example, the introduction of human stem cells into mice has been used to validate and characterize different types of stem cells. Some scientists hope the human cells will develop into specific tissues or organs for potential transplant purposes, or for drug testing. Other researchers are exploring the developmental nature of particular types of human cells, which may yield important insight into human biology and disease development. Animal models with human cells in the brain can be used to study many human brain diseases, including Parkinson’s, Alzheimer’s, and schizophrenia, and may be useful models for testing new drugs. Furthermore, these animal models could be used to study whether introduction of therapeutic human cells can improve outcomes for diseases that are caused by the death or dysfunction of neural or other brain cells, prior to testing the cells in people. 2) What kind of research does NIH anticipate funding in the future? This will depend on the ideas of the research community, but if researchers are successful in the development of animal models with human tissues or organs, those animal models could have many research applications for studying disease development and testing new therapeutics, as well as a possible source of tissues or organs for transplant. 3) Does NIH currently have any policy in this research area? Yes. The NIH Guidelines for Human Stem Cell Research prohibit the introduction of human pluripotent cells into non-human primate blastocyst-stage embryos, and prohibit the breeding of animals where the introduction of human pluripotent cells may have led to human sperm or human eggs being produced in the animal. -

An Interesting Case of Chimera: Inconclusive BRCA Analysis Due To
Case Report iMedPub Journals 2018 www.imedpub.com Journal of Genomics & Gene Study Vol.1 No.1:3 An Interesting Case of Chimera: Inconclusive Bowers AW*, Monteverde A, BRCA Analysis due to Two Distinct Genetic Keckeisen G and Kapenhas E Profiles in a Single Patient Ellen Hermanson Breast Center, Southampton, New York, USA Abstract *Corresponding author: Anne Wills Bowers Background: Our case study proposes an explanation for the inconclusive BRCA testing we received on one of our patients. The BRCA analysis was inconclusive [email protected] due to the specimen being chimeric, or having two distinct genetic profiles. We propose the Vanishing Twin theory as the most probable explanation. We have Ellen Hermanson Breast Center, found no other reported cases of chimera linked to Vanishing Twin Syndrome in Southampton, New York, USA. the literature. Methods: In this case report, we have described the presentation and operative Tel: +1 631-726-8400 course of a single patient of our Breast Center in 2017 to help define the setting of our unique BRCA analysis result. Findings: We have also done an extensive literature search in an attempt to Citation: Bowers AW, Monteverde A, understand the implications of the BRCA analysis result in a chimeric patient. Keckeisen G, Kapenhas E (2018) An However, our search was not satisfied by any direct evidence. We can only Interesting Case of Chimera: Inconclusive conclude that the Vanishing Twin theory is the most likely explanation of chimera BRCA Analysis due to Two Distinct Genetic in this single patient. Profiles in a Single Patient. J Genom Gene Study Vol.1 No.1:3 Conclusions: We were unable to find any direct explanation for our results but we do propose this as an interesting case, as chimera is rare. -
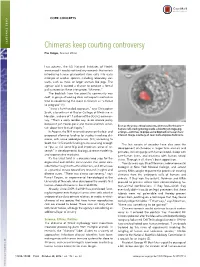
Chimeras Keep Courting Controversy Elie Dolgin, Science Writer
CORE CONCEPTS CORE CONCEPTS Chimeras keep courting controversy Elie Dolgin, Science Writer Last autumn, the US National Institutes of Health announced it would not fund any research that entails introducing human pluripotent stem cells into early embryos of another species, including laboratory stal- warts, such as mice, or larger animals like pigs. The agency said it wanted a chance to conduct a formal policy review on these interspecies “chimeras.” The backlash from the scientific community was swift. A group of leading stem cell experts wasted no time in condemning the move in Science as “a threat to progress” (1). “It was a ham-handed approach,” says Christopher Scott, a bioethicist at Baylor College of Medicine in Houston, and one of 11 authors of the Science commen- tary. “That’s a really terrible way to do science policy because it just creates panic and makes scientists uncer- — ” Even as they raise ethical concerns, chimeras like this one tain about their lines of inquiry. human cells (red) growing inside a blastocyst-stage pig In August, the NIH reversed course on the ban and embryo—continue to pique some biomedical researchers’ proposed allowing funding for studies involving chi- interest. Image courtesy of Juan Carlos Izpisua ´ Belmonte. meras, with some added provisos. Still, according to Scott, the 10.5-month funding hiatus was long enough The last couple of decades have also seen the “ to put at risk some big and important areas of re- development of chimeras in larger farm animals and ” search in developmental biology, disease modeling, primates, including pigs with human blood, sheep with and regenerative medicine.