Bird and Bat Predation Services in Tropical Forests and Agroforestry Landscapes
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Strategy Habitat: Ponderosa Pine Woodlands
Habitat: Conservation Summaries for Strategy Habitats Strategy Habitat: Ponderosa Pine Woodlands Ecoregions: Conservation Overview: Ponderosa Pine Woodlands are a Strategy Habitat in the Blue Moun- Ponderosa pine habitats historically covered a large portion of the tains, East Cascades, and Klamath Mountains ecoregions. Blue Mountains ecoregion, as well as parts of the East Cascades and Klamath Mountains. Ponderosa pine is still widely distributed in eastern Characteristics: and southern Oregon. However, the structure and species composition The structure and composition of ponderosa pine woodlands varies of woodlands have changed dramatically. Historically, ponderosa pine across the state, depending on local climate, soil type and moisture, habitats had frequent low-intensity ground fires that maintained an elevation, aspect and fire history. In Blue Mountains, East Cascades open understory. Due to past selective logging and fire suppression, and Klamath Mountains ecoregions, ponderosa pine woodlands have dense patches of smaller conifers have grown in the understory of pon- open canopies, generally covering 10-40 percent of the sky. Their derosa pine forests. Depending on the area, these conifers may include understories are variable combinations of shrubs, herbaceous plants, shade-tolerant Douglas-fir, grand fir and white fir, or young ponderosa and grasses. Ponderosa woodlands are dominated by ponderosa pine, pine and lodgepole pine. These dense stands are vulnerable to drought but may also have lodgepole, western juniper, aspen, western larch, stress, insect outbreaks, and disease. The tree layers act as ladder fuels, grand fir, Douglas-fir, incense cedar, sugar pine, or white fir, depend- increasing the chances that a ground fire will become a forest-destroy- ing on ecoregion and site conditions. -

North Cascades Contested Terrain
North Cascades NP: Contested Terrain: North Cascades National Park Service Complex: An Administrative History NORTH CASCADES Contested Terrain North Cascades National Park Service Complex: An Administrative History CONTESTED TERRAIN: North Cascades National Park Service Complex, Washington An Administrative History By David Louter 1998 National Park Service Seattle, Washington TABLE OF CONTENTS adhi/index.htm Last Updated: 14-Apr-1999 http://www.nps.gov/history/history/online_books/noca/adhi/[11/22/2013 1:57:33 PM] North Cascades NP: Contested Terrain: North Cascades National Park Service Complex: An Administrative History (Table of Contents) NORTH CASCADES Contested Terrain North Cascades National Park Service Complex: An Administrative History TABLE OF CONTENTS Cover Cover: The Southern Pickett Range, 1963. (Courtesy of North Cascades National Park) Introduction Part I A Wilderness Park (1890s to 1968) Chapter 1 Contested Terrain: The Establishment of North Cascades National Park Part II The Making of a New Park (1968 to 1978) Chapter 2 Administration Chapter 3 Visitor Use and Development Chapter 4 Concessions Chapter 5 Wilderness Proposals and Backcountry Management Chapter 6 Research and Resource Management Chapter 7 Dam Dilemma: North Cascades National Park and the High Ross Dam Controversy Chapter 8 Stehekin: Land of Freedom and Want Part III The Wilderness Park Ideal and the Challenge of Traditional Park Management (1978 to 1998) Chapter 9 Administration Chapter 10 http://www.nps.gov/history/history/online_books/noca/adhi/contents.htm[11/22/2013 -

Nezara Viridula L.) in Western Part of Romania
Trophic Evolution of Southern Green Stink Bugs (Nezara viridula l.) in Western Part of Romania 1 1 1 1 1 Ioana GROZEA2 *, Ana Maria1 VIRTEIU , Ramona STEF , Alin CARABET , Levente MOLNAR , Teodora FLORIAN and Mihai VLAD 1 Department of Biology and Plant Protection, Banat’s University of Agricultural Sciences and Veterinary2 Medicine “King Michael I of Romania” Timisoara, Romania. *)Department of Environment and Plant Protection, University of Agricultural Science and Medicine Veterinary Cluj Napoca corresponding author, e-mail: [email protected] BulletinUASVM Horticulture 72(2) / 2015 Print ISSN 1843-5254, Electronic ISSN 1843-5394 DOI:10.15835/buasvmcn-hort:11391 Abstract Nezara viridula L. (Hemiptera:Pentatomidae) is a relative a invasive species, highly poliphagous, species of solanaceous, legumes, cruciferous, malvaceous, grasses and other plants are affected. Considering that the insect which make the subject of this scientifically paper is a species detected in our country but unregistered and un- localized as a dangerous species we want to bring to attention potential spread and adaptation to new plant species, especially legumes and ornamental plants. Observations were made in Timis countyN. viridula in the western part of Romania (Timis), during the years 2013 and 2014. It was taken under observation by three types of plants (tomato crop, green space and fruit tree plantation). TheSyringa observations vulgaris Loniceraon plants japonica, have shown Hibiscus that sp. and Magnolia affected liliiflora all four. varieties of tomatoes, especiallyPrunus on fruits,persica both and greenPrunus and persica mature var. stage. nucipersica In green made space in only adults and nymphs was observed on follow ornamental plants: N. -

Territory Size and Stability in a Sedentary Neotropical Passerine: Is Resource Partitioning a Necessary Condition?
J. Field Ornithol. 76(4):395±401, 2005 Territory size and stability in a sedentary neotropical passerine: is resource partitioning a necessary condition? Janet V. Gorrell,1 Gary Ritchison,1,3 and Eugene S. Morton2 1 Department of Biological Sciences, Eastern Kentucky University, Richmond, Kentucky 40475 USA 2 Hemlock Hill Field Station, 22318 Teepleville Flats Road, Cambridge Springs, Pennsylvania 16403 USA Received 9 April 2004; accepted 11 April 2005 ABSTRACT. Long-term pair bonds and defense of territories year-round are common among tropical passer- ines. The boundaries of these territories tend to be stable, perhaps re¯ecting the need to defend an area that, regardless of conditions, provides suf®cient food resources. If, however, these stable territories are not, even tem- porarily, suf®ciently large, then intra-pair competition for available food may result, particularly in species with no sexual size dimorphism. With such competition, sex-speci®c differences in foraging behavior may result. Male and female Dusky Antbirds (Cercomacra tyrannina) are not size dimorphic, and pairs jointly defend territories throughout the year. Our objective was to determine if paired Dusky Antbirds exhibited sex-speci®c differences in foraging behavior. Foraging antbirds were observed in central Panama from February±July 2002 to determine if pairs par- titioned food resources. Males and females exhibited no differences in foraging behavior, with individuals of both sexes foraging at similar heights and using the same foraging maneuvers (glean, probe, and sally) and substrates (leaves, rolled leaves, and woody surfaces). These results suggest that Dusky Antbirds do not partition resources and that territory switching, rather than resource partitioning, may be the means by which they gain access to additional food resources. -

New Insights Into the Microbiota of Moth Pests
International Journal of Molecular Sciences Review New Insights into the Microbiota of Moth Pests Valeria Mereghetti, Bessem Chouaia and Matteo Montagna * ID Dipartimento di Scienze Agrarie e Ambientali, Università degli Studi di Milano, 20122 Milan, Italy; [email protected] (V.M.); [email protected] (B.C.) * Correspondence: [email protected]; Tel.: +39-02-5031-6782 Received: 31 August 2017; Accepted: 14 November 2017; Published: 18 November 2017 Abstract: In recent years, next generation sequencing (NGS) technologies have helped to improve our understanding of the bacterial communities associated with insects, shedding light on their wide taxonomic and functional diversity. To date, little is known about the microbiota of lepidopterans, which includes some of the most damaging agricultural and forest pests worldwide. Studying their microbiota could help us better understand their ecology and offer insights into developing new pest control strategies. In this paper, we review the literature pertaining to the microbiota of lepidopterans with a focus on pests, and highlight potential recurrent patterns regarding microbiota structure and composition. Keywords: symbiosis; bacterial communities; crop pests; forest pests; Lepidoptera; next generation sequencing (NGS) technologies; diet; developmental stages 1. Introduction Insects represent the most successful taxa of eukaryotic life, being able to colonize almost all environments, including Antarctica, which is populated by some species of chironomids (e.g., Belgica antarctica, Eretmoptera murphyi, and Parochlus steinenii)[1,2]. Many insects are beneficial to plants, playing important roles in seed dispersal, pollination, and plant defense (by feeding upon herbivores, for example) [3]. On the other hand, there are also damaging insects that feed on crops, forest and ornamental plants, or stored products, and, for these reasons, are they considered pests. -

1922 Elizabeth T
co.rYRIG HT, 192' The Moootainetro !scot1oror,d The MOUNTAINEER VOLUME FIFTEEN Number One D EC E M BER 15, 1 9 2 2 ffiount Adams, ffiount St. Helens and the (!oat Rocks I ncoq)Ora,tecl 1913 Organized 190!i EDITORlAL ST AitF 1922 Elizabeth T. Kirk,vood, Eclttor Margaret W. Hazard, Associate Editor· Fairman B. L�e, Publication Manager Arthur L. Loveless Effie L. Chapman Subsc1·iption Price. $2.00 per year. Annual ·(onl�') Se,·ent�·-Five Cents. Published by The Mountaineers lncorJ,orated Seattle, Washington Enlerecl as second-class matter December 15, 19t0. at the Post Office . at . eattle, "\Yash., under the .-\0t of March 3. 1879. .... I MOUNT ADAMS lllobcl Furrs AND REFLEC'rION POOL .. <§rtttings from Aristibes (. Jhoutribes Author of "ll3ith the <6obs on lltount ®l!!mµus" �. • � J� �·,,. ., .. e,..:,L....._d.L.. F_,,,.... cL.. ��-_, _..__ f.. pt",- 1-� r�._ '-';a_ ..ll.-�· t'� 1- tt.. �ti.. ..._.._....L- -.L.--e-- a';. ��c..L. 41- �. C4v(, � � �·,,-- �JL.,�f w/U. J/,--«---fi:( -A- -tr·�� �, : 'JJ! -, Y .,..._, e� .,...,____,� � � t-..__., ,..._ -u..,·,- .,..,_, ;-:.. � --r J /-e,-i L,J i-.,( '"'; 1..........,.- e..r- ,';z__ /-t.-.--,r� ;.,-.,.....__ � � ..-...,.,-<. ,.,.f--· :tL. ��- ''F.....- ,',L � .,.__ � 'f- f-� --"- ��7 � �. � �;')'... f ><- -a.c__ c/ � r v-f'.fl,'7'71.. I /!,,-e..-,K-// ,l...,"4/YL... t:l,._ c.J.� J..,_-...A 'f ',y-r/� �- lL.. ��•-/IC,/ ,V l j I '/ ;· , CONTENTS i Page Greetings .......................................................................tlristicles }!}, Phoiitricles ........ r The Mount Adams, Mount St. Helens, and the Goat Rocks Outing .......................................... B1/.ith Page Bennett 9 1 Selected References from Preceding Mount Adams and Mount St. -

The All-Bird Bulletin
Advancing Integrated Bird Conservation in North America Spring 2014 Inside this issue: The All-Bird Bulletin Protecting Habitat for 4 the Buff-breasted Sandpiper in Bolivia The Neotropical Migratory Bird Conservation Conserving the “Jewels 6 Act (NMBCA): Thirteen Years of Hemispheric in the Crown” for Neotropical Migrants Bird Conservation Guy Foulks, Program Coordinator, Division of Bird Habitat Conservation, U.S. Fish and Bird Conservation in 8 Wildlife Service (USFWS) Costa Rica’s Agricultural Matrix In 2000, responding to alarming declines in many Neotropical migratory bird popu- Uruguayan Rice Fields 10 lations due to habitat loss and degradation, Congress passed the Neotropical Migra- as Wintering Habitat for tory Bird Conservation Act (NMBCA). The legislation created a unique funding Neotropical Shorebirds source to foster the cooperative conservation needed to sustain these species through all stages of their life cycles, which occur throughout the Western Hemi- Conserving Antigua’s 12 sphere. Since its first year of appropriations in 2002, the NMBCA has become in- Most Critical Bird strumental to migratory bird conservation Habitat in the Americas. Neotropical Migratory 14 Bird Conservation in the The mission of the North American Bird Heart of South America Conservation Initiative is to ensure that populations and habitats of North Ameri- Aros/Yaqui River Habi- 16 ca's birds are protected, restored, and en- tat Conservation hanced through coordinated efforts at in- ternational, national, regional, and local Strategic Conservation 18 levels, guided by sound science and effec- in the Appalachians of tive management. The NMBCA’s mission Southern Quebec is to achieve just this for over 380 Neo- tropical migratory bird species by provid- ...and more! Cerulean Warbler, a Neotropical migrant, is a ing conservation support within and be- USFWS Bird of Conservation Concern and listed as yond North America—to Latin America Vulnerable on the International Union for Conser- Coordination and editorial vation of Nature (IUCN) Red List. -

Foraging Niche Structure and Coexistence in a Highly Diverse Community Of
Foraging niche structure and coexistence in a highly diverse community of Amazonian antbirds (Thamnophilidae: Aves) by SANTIAGO DAVID B.Sc, Universidad de Antioquia, 2011 A THESIS SUBMITTED IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF MASTER OF SCIENCE in THE FACULTY OF GRADUATE AND POSTDOCTORAL STUDIES (Zoology) THE UNIVERSITY OF BRITISH COLUMBIA (Vancouver) May 2016 © Santiago David, 2016 Abstract The high species richness of antbirds (Thamnophilidae) in Amazonian lowlands, where as many as 40 species may coexist at local scales, represents a major challenge for ecologists to explain patterns of coexistence and niche evolution. I studied the foraging ecology of a local community of antbirds in a 2-Km2 area of lowland forest in SE Peru to examine how co- occurring antbird species differ in their use of foraging resources, and whether these differences result in niche partitioning at the community level. I also examined whether resource use similarity was related to phylogenetic similarity in this local assemblage. Forty-four species of antbirds were detected in the study plot in a four-year period, with 30 species categorized as the local assemblage of common resident breeders. Multivariate analysis of foraging parameters showed that segregation at two height layers in two types of forest was more important than foraging substrates and maneuvers in explaining the differences observed in foraging behavior. However, a null model analysis revealed that at the community level, antbirds exhibited high foraging niche overlap, with average observed overlap significantly larger than expected by chance, indicating that antbirds prefer, rather than avoid, resources used by other species. -

Pine-Oak-Hemlock--Silviculture Institute 7/18/2017 1
Pine‐Oak‐Hemlock‐‐Silviculture Institute 7/18/2017 Predicted Responses of Wildlife to Silvicultural Treatments with special focus on Pine‐Oak‐Hemlock Types Matt Tarr Associate Extension Professor, Wildlife Specialist University of New Hampshire Cooperative Extension Northeast Silviculture Institute for Foresters June 18, 2017 What am I talking about? OUTLINE • Overview of the primary factors that determine how wildlife respond to timber harvesting • Overview of how birds, mammals, reptiles & amphibians select their habitat • Summary of expected wildlife response following application of silvicultural treatments in pine‐oak‐hemlock MY GOAL: Help you become better at predicting how wildlife will respond to the decisions you make when managing forested habitats 1 Pine‐Oak‐Hemlock‐‐Silviculture Institute 7/18/2017 When we cut trees, the response by wildlife is determined primarily by: • Plant species • Plant structure composition following the harvest • The size of the canopy opening • Whether soils are • Presence/absence of • Habitats in the dry/moist/wet special habitat features surrounding landscape “Plant structure” = presence & density of plants in each forest layer • What plant layers are present? Canopy layer >30’ Midstory layer 10-30’ Shrub layer 2-10’ Herbaceous layer 0-2’ • How dense are the plants in each layer? SPARSE INTERMEDIATE ABUNDANT/DENSE < 30% coverage 30-75% coverage > 75% coverage 2 Pine‐Oak‐Hemlock‐‐Silviculture Institute 7/18/2017 Role of plant structure in influencing wildlife use of forest stands Forest stands comprised -

Ecoregions of the Mississippi Alluvial Plain
92° 91° 90° 89° 88° Ecoregions of the Mississippi Alluvial Plain Cape Girardeau 73cc 72 io Ri Ecoregions denote areas of general similarity in ecosystems and in the type, quality, and quantity of This level III and IV ecoregion map was compiled at a scale of 1:250,000 and depicts revisions and Literature Cited: PRINCIPAL AUTHORS: Shannen S. Chapman (Dynamac Corporation), Oh ver environmental resources; they are designed to serve as a spatial framework for the research, subdivisions of earlier level III ecoregions that were originally compiled at a smaller scale (USEPA Bailey, R.G., Avers, P.E., King, T., and McNab, W.H., eds., 1994, Omernik, J.M., 1987, Ecoregions of the conterminous United States (map Barbara A. Kleiss (USACE, ERDC -Waterways Experiment Station), James M. ILLINOIS assessment, management, and monitoring of ecosystems and ecosystem components. By recognizing 2003, Omernik, 1987). This poster is part of a collaborative effort primarily between USEPA Region Ecoregions and subregions of the United States (map) (supplementary supplement): Annals of the Association of American Geographers, v. 77, no. 1, Omernik, (USEPA, retired), Thomas L. Foti (Arkansas Natural Heritage p. 118-125, scale 1:7,500,000. 71 the spatial differences in the capacities and potentials of ecosystems, ecoregions stratify the VII, USEPA National Health and Environmental Effects Research Laboratory (Corvallis, Oregon), table of map unit descriptions compiled and edited by McNab, W.H., and Commission), and Elizabeth O. Murray (Arkansas Multi-Agency Wetland Bailey, R.G.): Washington, D.C., U.S. Department of Agriculture - Forest Planning Team). 37° environment by its probable response to disturbance (Bryce and others, 1999). -
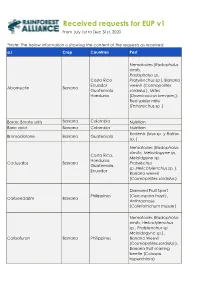
IIS 2020 Requests Overview
Received requests for EUP v1 From July 1st to Dec 31st, 2020 *Note: The below information is showing the content of the requests as received. a.i Crop Countries Pest Nematodes ( Radophulus similis, Radopholus sp, Costa Rica Pratylenchus sp ), Banana Ecuador weevil ( Cosmopolites Abamectin Banana Guatemala sordidus ), Mites Honduras (Dysmicoccus brevipes ), Red spider mite (Tetranychus sp .) Borax; Borate salts Banana Colombia Nutrition Boric acid Banana Colombia Nutrition Rodents ( Mus sp. y Rattus Bromadiolone Banana Guatemala sp. ) Nematodes ( Radopholus silmillis, Meloidogyne sp, Costa Rica, Meloidgune sp, Honduras, Cadusafos Banana Pratylechus Guatemala, sp.,Helicotylenchus sp. ), Ecuador Banana weevil (Cosmopolites sordidus ) Diamond Fruit Sport Philippines (Cercospora hayii ), Carbendazim Banana Anthracnose (Colletotrichum musae ) Nematod es (Radopholus similis, Helicotylenchus sp., Pratylenchus sp. Meloidogyne sp.) , Carbofuran Banana Philippines Banana Weevil (Cosmopolites sordidus ), Banana fruit scarring beetle (Colaspis hyperchlora) a.i Crop Countries Pest Black Sigatoka Ecuador, Costa (Mycosphaerella fijiensis ), Rica, Philippines, Yellow Sigatoka Chlorothalonil Banana Honduras, (Mycosphaerella Guatemala, musicola ), Banana Colombia Freckle ( Phyllosticta musarum ) Mealybug ( Dysmicoccus brevipes, Pseudococcus elisae, Pseudococcus sp, Ferrista sp ), Dysmicoccus sp , Scale insects (Aspidiotus destructor, Diaspis boisduvalii ), Thrips (Thrips florum, Franckliniella spp, Chaetanaphothrips signipennis ), Banana fruit Philippines, -

25Th U.S. Department of Agriculture Interagency Research Forum On
US Department of Agriculture Forest FHTET- 2014-01 Service December 2014 On the cover Vincent D’Amico for providing the cover artwork, “…and uphill both ways” CAUTION: PESTICIDES Pesticide Precautionary Statement This publication reports research involving pesticides. It does not contain recommendations for their use, nor does it imply that the uses discussed here have been registered. All uses of pesticides must be registered by appropriate State and/or Federal agencies before they can be recommended. CAUTION: Pesticides can be injurious to humans, domestic animals, desirable plants, and fish or other wildlife--if they are not handled or applied properly. Use all pesticides selectively and carefully. Follow recommended practices for the disposal of surplus pesticides and pesticide containers. Product Disclaimer Reference herein to any specific commercial products, processes, or service by trade name, trademark, manufacturer, or otherwise does not constitute or imply its endorsement, recom- mendation, or favoring by the United States government. The views and opinions of wuthors expressed herein do not necessarily reflect those of the United States government, and shall not be used for advertising or product endorsement purposes. The U.S. Department of Agriculture (USDA) prohibits discrimination in all its programs and activities on the basis of race, color, national origin, sex, religion, age, disability, political beliefs, sexual orientation, or marital or family status. (Not all prohibited bases apply to all programs.) Persons with disabilities who require alternative means for communication of program information (Braille, large print, audiotape, etc.) should contact USDA’s TARGET Center at 202-720-2600 (voice and TDD). To file a complaint of discrimination, write USDA, Director, Office of Civil Rights, Room 326-W, Whitten Building, 1400 Independence Avenue, SW, Washington, D.C.