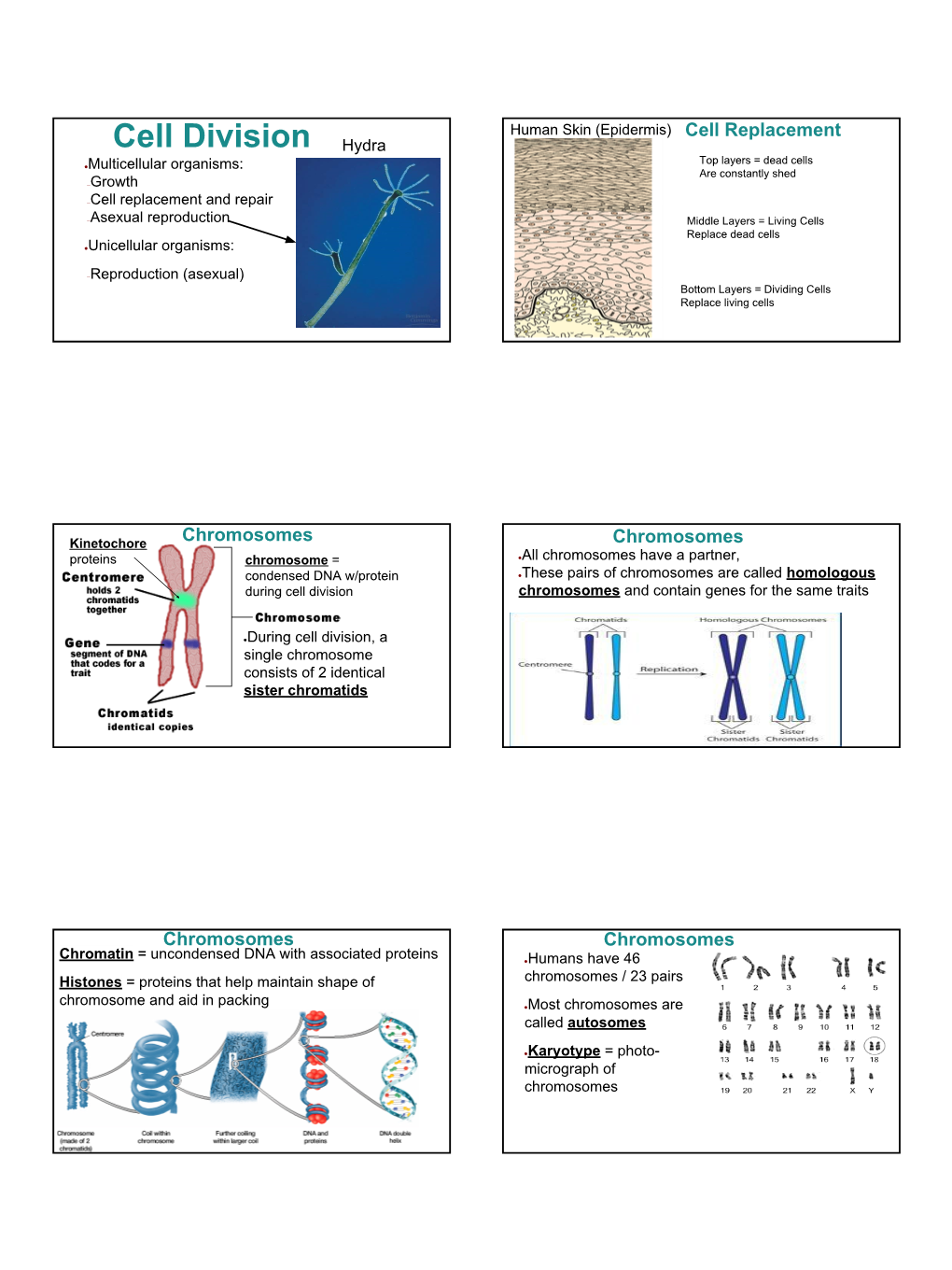
Cell Division Hydra Top Layers = Dead Cells ●Multicellular Organisms: Are Constantly Shed –Growth
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

RASSF1A Interacts with and Activates the Mitotic Kinase Aurora-A
Oncogene (2008) 27, 6175–6186 & 2008 Macmillan Publishers Limited All rights reserved 0950-9232/08 $32.00 www.nature.com/onc ORIGINAL ARTICLE RASSF1A interacts with and activates the mitotic kinase Aurora-A L Liu1, C Guo1, R Dammann2, S Tommasi1 and GP Pfeifer1 1Division of Biology, Beckman Research Institute, City of Hope Cancer Center, Duarte, CA, USA and 2Institute of Genetics, University of Giessen, Giessen, Germany The RAS association domain family 1A (RASSF1A) gene tumorigenesis and carcinogen-induced tumorigenesis is located at chromosome 3p21.3 within a specific area of (Tommasi et al., 2005; van der Weyden et al., 2005), common heterozygous and homozygous deletions. RASS- supporting the notion that RASSF1A is a bona fide F1A frequently undergoes promoter methylation-asso- tumor suppressor. However, it is not fully understood ciated inactivation in human cancers. Rassf1aÀ/À mice how RASSF1A is involved in tumor suppression. are prone to both spontaneous and carcinogen-induced The biochemical function of the RASSF1A protein is tumorigenesis, supporting the notion that RASSF1A is a largely unknown. The homology of RASSF1A with the tumor suppressor. However, it is not fully understood how mammalian Ras effector novel Ras effector (NORE)1 RASSF1A is involved in tumor suppression pathways. suggests that the RASSF1A gene product may function Here we show that overexpression of RASSF1A inhibits in signal transduction pathways involving Ras-like centrosome separation. RASSF1A interacts with Aurora-A, proteins. However, recent data indicate that RASSF1A a mitotic kinase. Surprisingly, knockdown of RASS- itself binds to RAS only weakly and that binding to F1A by siRNA led to reduced activation of Aurora-A, RAS may require heterodimerization of RASSF1A and whereas overexpression of RASSF1A resulted in in- NORE1 (Ortiz-Vega et al., 2002). -

Mitosis Vs. Meiosis
Mitosis vs. Meiosis In order for organisms to continue growing and/or replace cells that are dead or beyond repair, cells must replicate, or make identical copies of themselves. In order to do this and maintain the proper number of chromosomes, the cells of eukaryotes must undergo mitosis to divide up their DNA. The dividing of the DNA ensures that both the “old” cell (parent cell) and the “new” cells (daughter cells) have the same genetic makeup and both will be diploid, or containing the same number of chromosomes as the parent cell. For reproduction of an organism to occur, the original parent cell will undergo Meiosis to create 4 new daughter cells with a slightly different genetic makeup in order to ensure genetic diversity when fertilization occurs. The four daughter cells will be haploid, or containing half the number of chromosomes as the parent cell. The difference between the two processes is that mitosis occurs in non-reproductive cells, or somatic cells, and meiosis occurs in the cells that participate in sexual reproduction, or germ cells. The Somatic Cell Cycle (Mitosis) The somatic cell cycle consists of 3 phases: interphase, m phase, and cytokinesis. 1. Interphase: Interphase is considered the non-dividing phase of the cell cycle. It is not a part of the actual process of mitosis, but it readies the cell for mitosis. It is made up of 3 sub-phases: • G1 Phase: In G1, the cell is growing. In most organisms, the majority of the cell’s life span is spent in G1. • S Phase: In each human somatic cell, there are 23 pairs of chromosomes; one chromosome comes from the mother and one comes from the father. -

Cell Division- Ch 5
Cell Division- Mitosis and Meiosis When do cells divide? Cell size . One of most important factors affecting size of the cell is size of cell membrane . Cell must remain relatively small to survive (why?) – Cell membrane has to be big enough to take in nutrients and eliminate wastes – As cells get bigger, the volume increases faster than the surface area – Small cells have a larger surface area to volume ratio than larger cells to help with nutrient intake and waste elimination . When a cell reaches its max size, the nucleus starts cell division: called MITOSIS or MEIOSIS Mitosis . General Information – Occurs in somatic (body) cells ONLY!! – Nickname: called “normal” cell division – Produces somatic cells only . Background Info – Starts with somatic cell in DIPLOID (2n) state . Cell contains homologous chromosomes- chromosomes that control the same traits but not necessarily in the same way . 1 set from mom and 1 set from dad – Ends in diploid (2n) state as SOMATIC cells – Goes through one set of divisions – Start with 1 cell and end with 2 cells Mitosis (cont.) . Accounts for three essential life processes – Growth . Result of cell producing new cells . Develop specialized shapes/functions in a process called differentiation . Rate of cell division controlled by GH (Growth Hormone) which is produced in the pituitary gland . Ex. Nerve cell, intestinal cell, etc. – Repair . Cell regenerates at the site of injury . Ex. Skin (replaced every 28 days), blood vessels, bone Mitosis (cont.) – Reproduction . Asexual – Offspring produced by only one parent – Produce offspring that are genetically identical – MITOSIS – Ex. Bacteria, fungi, certain plants and animals . -

The Kinase Activity of Aurora B Is Required for Kinetochore-Microtubule Interactions During Mitosis
Current Biology, Vol. 12, 894–899, June 4, 2002, 2002 Elsevier Science Ltd. All rights reserved. PII S0960-9822(02)00848-5 The Kinase Activity of Aurora B Is Required for Kinetochore-Microtubule Interactions during Mitosis Maki Murata-Hori and Yu-li Wang1 chromosomal congression, we monitored the move- Department of Physiology ment of individual centromeres in cells expressing a high University of Massachusetts Medical School level of aurora B(K-R)-GFP or control cells expressing a Worcester, Massachusetts 01605 similar level of wild-type aurora B-GFP (Figure 1B). Both aurora B-GFP and aurora B(K-R)-GFP were localized at centromeres and spindle poles [6] and to a less extent Summary along chromosomal arms (see Figures 3A and 3D). In control cells, the centromeres of neighboring chro- As a component of the “chromosomal passenger pro- mosomes moved independently of each other at an av- Ϯ Ϯ ϭ tein complex,” the aurora B kinase is associated with erage rate of 1.8 1.2 m/min (mean SD, n 22), centromeres during prometaphase and with midzone with frequent changes in direction (Figure 1B, left). They ف microtubules during anaphase and is required for both eventually accumulated at the metaphase plate 20 min mitosis and cytokinesis [1–6]. Ablation of aurora B after nuclear envelope breakdown (19/19, Figures 1Aa– causes defects in both prometaphase chromosomal 1Ad; Supplementary Movie 1 available with this article congression and the spindle checkpoint [4–6]; how- online). In contrast, in cells expressing aurora B(K-R)- ever, the mechanisms underlying these defects are GFP, centromeres of neighboring chromosomes moved Ϯ unclear. -

Ran Controls Microtubule Asters and Nuclear Assembly 2455 Chromatin Rounded up (Fig
Journal of Cell Science 112, 2453-2461 (1999) 2453 Printed in Great Britain © The Company of Biologists Limited 1999 JCS0524 Ran-GTP stabilises microtubule asters and inhibits nuclear assembly in Xenopus egg extracts Chuanmao Zhang1,2, Mike Hughes1 and Paul R. Clarke1,* 1Biomedical Research Centre, University of Dundee, Level 5, Ninewells Hospital and Medical School, Dundee DD1 9SY, Scotland, UK 2Department of Cell Biology and Genetics, College of Life Sciences, Peking University, Beijing 100871, China *Author for correspondence (e-mail: [email protected]) Accepted 25 May; published on WWW 24 June 1999 SUMMARY Ran is an abundant GTPase of the Ras superfamily that is nucleus and blocks chromatin decondensation. In contrast, highly conserved in eukaryotes. In interphase cells, Ran is Ran GDP does not stabilise microtubules or inhibit nuclear mainly nuclear and thought to be predominantly GTP- assembly. RanT24N and RanBP1, which oppose the bound, but it is also present in the cytoplasm, probably generation of Ran-GTP by RCC1, arrest nuclear growth GDP-bound. This asymmetric distribution plays an after disappearance of the aster. Ran associates with important role in directing nucleocytoplasmic transport. microtubule asters in egg extracts and with mitotic spindles Ran has also been implicated in cell cycle control, including in somatic Xenopus cells, suggesting that it may affect the transition from mitosis to interphase when the microtubule stability directly. These results show that Ran compartmentalisation of the nucleus is established. Here, has a novel function in the control of microtubule stability we have examined the role of Ran in this transition using that is clearly distinct from nucleocytoplasmic transport. -

Mitosin/CENP-F in Mitosis, Transcriptional Control, and Differentiation
Journal of Biomedical Science (2006) 13: 205–213 205 DOI 10.1007/s11373-005-9057-3 Mitosin/CENP-F in mitosis, transcriptional control, and differentiation Li Ma1,2, Xiangshan Zhao1 & Xueliang Zhu1,* 1Laboratory of Molecular Cell Biology, Institute of Biochemistry and Cell Biology, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences, Shanghai, 200031, China; 2Graduate School of Chinese Academy of Sciences, Beijing, 100039, China Ó 2006 National Science Council, Taipei Key words: differentiation, kinetochore, mitosis, transcription Summary Mitosin/CENP-F is a large nuclear/kinetochore protein containing multiple leucine zipper motifs poten- tially for protein interactions. Its expression levels and subcellular localization patterns are regulated in a cell cycle-dependent manner. Recently, accumulating lines of evidence have suggested it a multifunctional protein involved in mitotic control, microtubule dynamics, transcriptional regulation, and muscle cell differentiation. Consistently, it is shown to interact directly with a variety of proteins including CENP-E, NudE/Nudel, ATF4, and Rb. Here we review the current progress and discuss possible mechanisms through which mitosin may function. Mitosin, also named CENP-F, was initially iden- acid residues (GenBank Accession No. CAH73032) tified as a human autoimmune antigen [1, 2] and [3]. The gene expression is cell cycle-dependent, as an in vitro binding protein of the tumor suppressor the mRNA levels increase following S phase Rb [3, 4]. Its dynamic temporal expression and progression and peak in early M phase [3]. Such modification patterns as well as ever-changing patterns are regulated by the Forkhead transcrip- spatial distributions following the cell cycle pro- tion factor FoxM1 [5]. -

Kinetochores, Microtubules, and Spindle Assembly Checkpoint
Review Joined at the hip: kinetochores, microtubules, and spindle assembly checkpoint signaling 1 1,2,3 Carlos Sacristan and Geert J.P.L. Kops 1 Molecular Cancer Research, University Medical Center Utrecht, 3584 CG Utrecht, The Netherlands 2 Center for Molecular Medicine, University Medical Center Utrecht, 3584 CG Utrecht, The Netherlands 3 Cancer Genomics Netherlands, University Medical Center Utrecht, 3584 CG Utrecht, The Netherlands Error-free chromosome segregation relies on stable and cell division. The messenger is the SAC (also known as connections between kinetochores and spindle microtu- the mitotic checkpoint) (Figure 1). bules. The spindle assembly checkpoint (SAC) monitors The transition to anaphase is triggered by the E3 ubiqui- such connections and relays their absence to the cell tin ligase APC/C, which tags inhibitors of mitotic exit cycle machinery to delay cell division. The molecular (CYCLIN B) and of sister chromatid disjunction (SECURIN) network at kinetochores that is responsible for microtu- for proteasomal degradation [2]. The SAC has a one-track bule binding is integrated with the core components mind, inhibiting APC/C as long as incorrectly attached of the SAC signaling system. Molecular-mechanistic chromosomes persist. It goes about this in the most straight- understanding of how the SAC is coupled to the kineto- forward way possible: it assembles a direct and diffusible chore–microtubule interface has advanced significantly inhibitor of APC/C at kinetochores that are not connected in recent years. The latest insights not only provide a to spindle microtubules. This inhibitor is named the striking view of the dynamics and regulation of SAC mitotic checkpoint complex (MCC) (Figure 1). -

Kinetochore Structure, Duplication, and Distribution in Mammalian Cells : Analysis by Human Autoantibodies from Scleroderma Patients
Kinetochore Structure, Duplication, and Distribution in Mammalian Cells : Analysis by Human Autoantibodies from Scleroderma Patients SARI BRENNER, DANIEL PEPPER, M . W. BERNS, E . TAN, and B . R . BRINKLEY Department of Cell Biology, Baylor College of Medicine, Houston, Texas 77030, High Voltage Electron Microscope Laboratory, Madison, Wisconsin 53706, Department of Developmental and Cell Biology, University of California, Irvine, Irvine, California 92717, and Department of Medicine, University of Colorado, Medical Center, Denver, Colorado 80262 ABSTRACT The specificity of the staining of CREST scleroderma patient serum was investigated by immunofluorescence and immunoelectron microscopy. The serum was found to stain the centromere region of mitotic chromosomes in many mammalian cell types by immunofluores- cence. It also localized discrete spots in interphase nuclei which we have termed "presumptive kinetochores ." The number of presumptive kinetochores per cell corresponds to the chromo- some number in the cell lines observed . Use of the immunoperoxidase technique to localize the antisera on PtK2 cells at the electron microscopic level revealed the specificity of the sera for the trilaminar kinetochore disks on metaphase and anaphase chromosomes . Presumptive kinetochores in the interphase nuclei were also visible in the electron microscope as randomly arranged, darkly stained spheres averaging 0.22 p,m in diameter. Preabsorption of the antisera was attempted using microtubule protein, purified tubulin, actin, and microtubule-associated proteins . None of these proteins diminished the immunofluorescence staining of the sera, indicating that the antibody-specific antigen(s) is a previously unrecognized component of the kinetochore region . In some interphase cells observed by both immunofluorescence and immunoelectron microscopy, the presumptive kinetochores appeared as double rather than single spots . -

Molecular Biology and Applied Genetics
MOLECULAR BIOLOGY AND APPLIED GENETICS FOR Medical Laboratory Technology Students Upgraded Lecture Note Series Mohammed Awole Adem Jimma University MOLECULAR BIOLOGY AND APPLIED GENETICS For Medical Laboratory Technician Students Lecture Note Series Mohammed Awole Adem Upgraded - 2006 In collaboration with The Carter Center (EPHTI) and The Federal Democratic Republic of Ethiopia Ministry of Education and Ministry of Health Jimma University PREFACE The problem faced today in the learning and teaching of Applied Genetics and Molecular Biology for laboratory technologists in universities, colleges andhealth institutions primarily from the unavailability of textbooks that focus on the needs of Ethiopian students. This lecture note has been prepared with the primary aim of alleviating the problems encountered in the teaching of Medical Applied Genetics and Molecular Biology course and in minimizing discrepancies prevailing among the different teaching and training health institutions. It can also be used in teaching any introductory course on medical Applied Genetics and Molecular Biology and as a reference material. This lecture note is specifically designed for medical laboratory technologists, and includes only those areas of molecular cell biology and Applied Genetics relevant to degree-level understanding of modern laboratory technology. Since genetics is prerequisite course to molecular biology, the lecture note starts with Genetics i followed by Molecular Biology. It provides students with molecular background to enable them to understand and critically analyze recent advances in laboratory sciences. Finally, it contains a glossary, which summarizes important terminologies used in the text. Each chapter begins by specific learning objectives and at the end of each chapter review questions are also included. -

The Emerging Role of Ncrnas and RNA-Binding Proteins in Mitotic Apparatus Formation
non-coding RNA Review The Emerging Role of ncRNAs and RNA-Binding Proteins in Mitotic Apparatus Formation Kei K. Ito, Koki Watanabe and Daiju Kitagawa * Department of Physiological Chemistry, Graduate School of Pharmaceutical Science, The University of Tokyo, Bunkyo, Tokyo 113-0033, Japan; [email protected] (K.K.I.); [email protected] (K.W.) * Correspondence: [email protected] Received: 11 November 2019; Accepted: 13 March 2020; Published: 20 March 2020 Abstract: Mounting experimental evidence shows that non-coding RNAs (ncRNAs) serve a wide variety of biological functions. Recent studies suggest that a part of ncRNAs are critically important for supporting the structure of subcellular architectures. Here, we summarize the current literature demonstrating the role of ncRNAs and RNA-binding proteins in regulating the assembly of mitotic apparatus, especially focusing on centrosomes, kinetochores, and mitotic spindles. Keywords: ncRNA; centrosome; kinetochore; mitotic spindle 1. Introduction Non-coding RNAs (ncRNAs) are defined as a class of RNA molecules that are transcribed from genomic DNA, but not translated into proteins. They are mainly classified into the following two categories according to their length—small RNA (<200 nt) and long non-coding RNA (lncRNA) (>200 nt). Small RNAs include traditional RNA molecules, such as transfer RNA (tRNA), small nuclear RNA (snRNA), small nucleolar RNA (snoRNA), PIWI-interacting RNA (piRNA), and micro RNA (miRNA), and they have been studied extensively [1]. Research on lncRNA is behind that on small RNA despite that recent transcriptome analysis has revealed that more than 120,000 lncRNAs are generated from the human genome [2–4]. -

Interferon-A-Induced G1 Phase Arrest Through Up-Regulated Expression of CDK Inhibitors, P19ink4d and P21cip1 in Mouse Macrophages
Oncogene (1998) 16, 2075 ± 2086 1998 Stockton Press All rights reserved 0950 ± 9232/98 $12.00 http://www.stockton-press.co.uk/onc Interferon-a-induced G1 phase arrest through up-regulated expression of CDK inhibitors, p19Ink4D and p21Cip1 in mouse macrophages Masaaki Matsuoka, Kenzaburo Tani and Shigetaka Asano Department of Hematology and Oncology, Institute of Medical Science, University of Tokyo, 4-6-1 Shirokanedai, Minato-ku, Tokyo 108, Japan The mechanism of cell cycle arrest induced by interferon-a dependent kinases or cdks (reviewed by Sherr, 1993). (IFN-a) was analysed using a mouse macrophage cell line, D-type cyclins (Lew et al., 1991; Matsushime et al., 1991) BAC1.2F5A. IFN-a added in media before mid-G1 and cyclin E (Ko et al., 1991; Lew et al., 1991) govern the prohibited cells from entering S phase. The blockage of G1/S transition in association with their proper G1/S transition was associated with diminuition of both physiological partners, cdk4 (Matsushime et al., 1992) cyclin D1/cdk4- and cyclin E/cdk2-associated kinase or cdk6 (Meyerson and Harlow, 1994), and cdk2 (Dulic et activities. G1 cyclin-associated kinase activities were al., 1992; Ko et al., 1992), respectively. When cells enter down-regulated quickly after the addition of IFN-a. Cells G1 phase with mitogenic stimuli, the synthesis and the treated with IFN-a contained excess amounts of cdk active complex formation of D-type cyclins and cdk4 are inhibitors which down-regulated G1 cyclin/cdk-associated induced in early to middle G1 phase, and their complex kinase activities in the proliferating cells and this action formation and activities reach maximal levels at late G1 was counteracted by exogenously-supplied recombinant (Matsushime et al., 1992, 1994). -

Targeting the WEE1 Kinase As a Molecular Targeted Therapy for Gastric Cancer
www.impactjournals.com/oncotarget/ Oncotarget, Vol. 7, No. 31 Research Paper Targeting the WEE1 kinase as a molecular targeted therapy for gastric cancer Hye-Young Kim1,2, Yunhee Cho1,3, HyeokGu Kang1,3, Ye-Seal Yim1,3, Seok-Jun Kim1,3, Jaewhan Song2, Kyung-Hee Chun1,3 1Department of Biochemistry & Molecular Biology, Yonsei University College of Medicine, Seodaemun-gu, Seoul 03722, Korea 2Department of Biochemistry, College of Life Science and Biotechnology, Seodaemun-gu, Seoul 03722, Korea 3Brain Korea 21 PlusProject for Medical Science, Yonsei University, Seodaemun-gu, Seoul 03722, Korea Correspondence to: Kyung-Hee Chun, email: [email protected] Keywords: WEE1, AZD1775 (MK-1775), 5-FU, Paclitaxel, gastric cancer Received: September 07, 2015 Accepted: May 28, 2016 Published: June 23, 2016 ABSTRACT Wee1 is a member of the Serine/Threonine protein kinase family and is a key regulator of cell cycle progression. It has been known that WEE1 is highly expressed and has oncogenic functions in various cancers, but it is not yet studied in gastric cancers. In this study, we investigated the oncogenic role and therapeutic potency of targeting WEE1 in gastric cancer. At first, higher expression levels of WEE1 with lower survival probability were determined in stage 4 gastric cancer patients or male patients with accompanied lymph node metastasis. To determine the function of WEE1 in gastric cancer cells, we determined that WEE1 ablation decreased the proliferation, migration, and invasion, while overexpression of WEE1 increased these effects in gastric cancer cells. We also validated the clinical application of WEE1 targeting by a small molecule, AZD1775 (MK-1775), which is a WEE1 specific inhibitor undergoing clinical trials.