The Morphology of the Axillary Border of the Scapula with Special Reference to the Neandertal Problem
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-
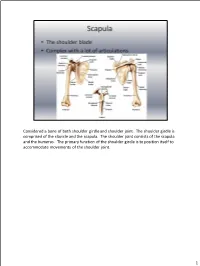
Considered a Bone of Both Shoulder Girdle and Shoulder Joint. the Shoulder Girdle Is Comprised of the Clavicle and the Scapula
Considered a bone of both shoulder girdle and shoulder joint. The shoulder girdle is comprised of the clavicle and the scapula. The shoulder joint consists of the scapula and the humerus. The primary function of the shoulder girdle is to position itself to accommodate movements of the shoulder joint. 1 Superior angle—top point Inferior angle—bottom point Vertebral border—side closest to vertebral column Axillary border—side closest to arm Subscapular fossa—anterior fossa Glenoid fossa, glenoid labrum, glenoid cavity --The glenoid fossa is the shallow cavity where the humeral head goes. The glenoid labrum is the cartilage that goes around the glenoid fossa. So the glenoid fossa and glenoid labrum together comprise the glenoid cavity. Supraspinous fossa—posterior, fossa above the spine Spine of the scapula—the back projection Infraspinous fossa—posterior depression/fossa below spine Coracoid process—anterior projection head Acromion process—posterior projection head above spine 2 Scapulothoracic “joint” = NOT a true joint; there are no ligaments or articular capsule. The scapula just rests on the muscle over top the rib cage, which allows for passive movements. Sternoclavicular joint=where the clavicle (collarbone) and the sternum (breastbone) articulate; movement is slight in all directions and of a gliding, rotational type Acromioclavicular joint = where the clavicle and scapula (acromion process) articulate; AKA: AC Joint; movement is a slight gliding when elevation and depression take place. Glenohumeral joint = the shoulder joint 3 4 All 3 true joints: Sternoclavicular, AC and glenohumeral (GH) all work together to move arm in all directions. The GH allows the arm to go out to the side and be abducted, then the AC and Sternoclavicular joints kick in to allow the arm to go above shoulder level by allowing the shoulderblade to move up to increase the range of motion (ROM). -

Scapular Motion Tracking Using Acromion Skin Marker Cluster: in Vitro Accuracy Assessment
Scapular Motion Tracking Using Acromion Skin Marker Cluster: In Vitro Accuracy Assessment Andrea Cereatti, Claudio Rosso, Ara Nazarian, Joseph P. DeAngelis, Arun J. Ramappa & Ugo Della Croce Journal of Medical and Biological Engineering ISSN 1609-0985 J. Med. Biol. Eng. DOI 10.1007/s40846-015-0010-2 1 23 Your article is protected by copyright and all rights are held exclusively by Taiwanese Society of Biomedical Engineering. This e- offprint is for personal use only and shall not be self-archived in electronic repositories. If you wish to self-archive your article, please use the accepted manuscript version for posting on your own website. You may further deposit the accepted manuscript version in any repository, provided it is only made publicly available 12 months after official publication or later and provided acknowledgement is given to the original source of publication and a link is inserted to the published article on Springer's website. The link must be accompanied by the following text: "The final publication is available at link.springer.com”. 1 23 Author's personal copy J. Med. Biol. Eng. DOI 10.1007/s40846-015-0010-2 ORIGINAL ARTICLE Scapular Motion Tracking Using Acromion Skin Marker Cluster: In Vitro Accuracy Assessment Andrea Cereatti • Claudio Rosso • Ara Nazarian • Joseph P. DeAngelis • Arun J. Ramappa • Ugo Della Croce Received: 11 October 2013 / Accepted: 20 March 2014 Ó Taiwanese Society of Biomedical Engineering 2015 Abstract Several studies have recently investigated how estimated using an AMC combined with a single anatom- the implementations of acromion marker clusters (AMCs) ical calibration, the accuracy was highly dependent on the method and stereo-photogrammetry affect the estimates of specimen and the type of motion (maximum errors between scapula kinematics. -

Phty 101 – Week 1
PHTY 101 – WEEK 1 1.7 Classify the shoulder (glenohumeral) joint and identify and/or describe its: • The glenohumeral joint is a multiaxial, synovial, ball and socket joint • Movements: o Flexion and extension- transverse axis o Abduction and adduction- anteroposterior axis o Internal and external rotation- longitudinal axis • Greatest amount of motion and most unstable joint in body • Function: positions and directs humerus, elbow and hand in relation to trunk (radioulnar joint deal with palm and fingers) • Combines with scapulothoracic, acromioclavicular and sternoclavicular joints • Articular surfaces: o Humeral head- ½ sphere, faces medially o Glenoid fossa- very shallow, faces laterally o Only 25-30% contact between articular surfaces o Shallower and smaller surface compared to hip • Glenoid labrum o Glenoid labrum- fibrous structure around glenoid fossa (adheres to edges), central part avascular and edges not greatly vascularised à doesn’t heal well, labrum functions to; - Facilitate mobility - Increase glenoid concavity- increasing mobility - Provide attachment for joint capsule, ligaments and muscles • Joint capsule o Very thin and lax- doesn’t compromise ROM o Attaches to glenoid labrum o Reflected (folded) inferiorly onto medial shaft of humerus o Reinforced by; - Rotator cuff tendons- supraspinatus, infraspinatus, teres minor, subscapularis - Rotator cuff tendons- action: internal or external rotation, function: pull humerus head in against fossa - Glenohumeral and coracohumeral ligaments (capsular) • Synovial membrane o Lines -

Bone Limb Upper
Shoulder Pectoral girdle (shoulder girdle) Scapula Acromioclavicular joint proximal end of Humerus Clavicle Sternoclavicular joint Bone: Upper limb - 1 Scapula Coracoid proc. 3 angles Superior Inferior Lateral 3 borders Lateral angle Medial Lateral Superior 2 surfaces 3 processes Posterior view: Acromion Right Scapula Spine Coracoid Bone: Upper limb - 2 Scapula 2 surfaces: Costal (Anterior), Posterior Posterior view: Costal (Anterior) view: Right Scapula Right Scapula Bone: Upper limb - 3 Scapula Glenoid cavity: Glenohumeral joint Lateral view: Infraglenoid tubercle Right Scapula Supraglenoid tubercle posterior anterior Bone: Upper limb - 4 Scapula Supraglenoid tubercle: long head of biceps Anterior view: brachii Right Scapula Bone: Upper limb - 5 Scapula Infraglenoid tubercle: long head of triceps brachii Anterior view: Right Scapula (with biceps brachii removed) Bone: Upper limb - 6 Posterior surface of Scapula, Right Acromion; Spine; Spinoglenoid notch Suprspinatous fossa, Infraspinatous fossa Bone: Upper limb - 7 Costal (Anterior) surface of Scapula, Right Subscapular fossa: Shallow concave surface for subscapularis Bone: Upper limb - 8 Superior border Coracoid process Suprascapular notch Suprascapular nerve Posterior view: Right Scapula Bone: Upper limb - 9 Acromial Clavicle end Sternal end S-shaped Acromial end: smaller, oval facet Sternal end: larger,quadrangular facet, with manubrium, 1st rib Conoid tubercle Trapezoid line Right Clavicle Bone: Upper limb - 10 Clavicle Conoid tubercle: inferior -
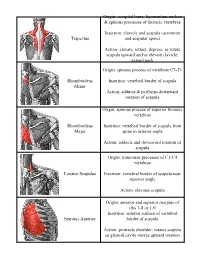
Trapezius Origin: Occipital Bone, Ligamentum Nuchae & Spinous Processes of Thoracic Vertebrae Insertion: Clavicle and Scapul
Origin: occipital bone, ligamentum nuchae & spinous processes of thoracic vertebrae Insertion: clavicle and scapula (acromion Trapezius and scapular spine) Action: elevate, retract, depress, or rotate scapula upward and/or elevate clavicle; extend neck Origin: spinous process of vertebrae C7-T1 Rhomboideus Insertion: vertebral border of scapula Minor Action: adducts & performs downward rotation of scapula Origin: spinous process of superior thoracic vertebrae Rhomboideus Insertion: vertebral border of scapula from Major spine to inferior angle Action: adducts and downward rotation of scapula Origin: transverse precesses of C1-C4 vertebrae Levator Scapulae Insertion: vertebral border of scapula near superior angle Action: elevates scapula Origin: anterior and superior margins of ribs 1-8 or 1-9 Insertion: anterior surface of vertebral Serratus Anterior border of scapula Action: protracts shoulder: rotates scapula so glenoid cavity moves upward rotation Origin: anterior surfaces and superior margins of ribs 3-5 Insertion: coracoid process of scapula Pectoralis Minor Action: depresses & protracts shoulder, rotates scapula (glenoid cavity rotates downward), elevates ribs Origin: supraspinous fossa of scapula Supraspinatus Insertion: greater tuberacle of humerus Action: abduction at the shoulder Origin: infraspinous fossa of scapula Infraspinatus Insertion: greater tubercle of humerus Action: lateral rotation at shoulder Origin: clavicle and scapula (acromion and adjacent scapular spine) Insertion: deltoid tuberosity of humerus Deltoid Action: -

Alliance Glenoid Surgical Technique
Alliance™ Glenoid Surgical Technique Table of Contents Indications/Contraindications ................................................................................. 2 Introduction .............................................................................................................. 3 Compatibility with Other Zimmer Biomet Humeral Heads ..................................... 3 Pre-Operative Planning ............................................................................................ 3 Technique Summary ................................................................................................. 4 4-Peg Modular Glenoid ............................................................................................ 9 4-Peg Modular Augmented Glenoid ...................................................................... 13 3-Peg Modular Glenoid .......................................................................................... 20 3-Peg Monoblock Glenoid ...................................................................................... 24 2-Peg Monoblock Glenoid ...................................................................................... 28 Revision ................................................................................................................... 32 2 | Alliance Glenoid Surgical Technique The Alliance Glenoid has the following indications/intended uses: INDICATIONS CONTRAINDICATIONS • Non-inflammatory degenerative joint disease Absolute contraindications include infection, sepsis, including -

The Role of Scapular Morphology in Reverse Shoulder Arthroplasty
Western University Scholarship@Western Electronic Thesis and Dissertation Repository 12-7-2016 12:00 AM The Role of Scapular Morphology in Reverse Shoulder Arthroplasty Ashish Gupta The University of Western Ontario Supervisor Dr Louis M Ferreira The University of Western Ontario Joint Supervisor Dr George Athwal The University of Western Ontario Graduate Program in Surgery A thesis submitted in partial fulfillment of the equirr ements for the degree in Master of Science © Ashish Gupta 2016 Follow this and additional works at: https://ir.lib.uwo.ca/etd Part of the Biomechanics and Biotransport Commons, Biomedical Devices and Instrumentation Commons, and the Orthopedics Commons Recommended Citation Gupta, Ashish, "The Role of Scapular Morphology in Reverse Shoulder Arthroplasty" (2016). Electronic Thesis and Dissertation Repository. 4357. https://ir.lib.uwo.ca/etd/4357 This Dissertation/Thesis is brought to you for free and open access by Scholarship@Western. It has been accepted for inclusion in Electronic Thesis and Dissertation Repository by an authorized administrator of Scholarship@Western. For more information, please contact [email protected]. Abstract Baseplate fixation in a reverse shoulder arthroplasty depends on adequate bone stock. In cases of severe glenoid bone loss and revision shoulder arthroplasty, deficiency of the glenoid vault compels the surgeon to attain screw fixation in the three columns of the scapula. The relationship of these columns demonstrated that the coracoid is closer to the lateral scapular pillar in females than in males. Significant gender dimorphism exists between the orientations of the three columns. The gender dimorphism is further evaluated by anthropometric measurements of the scapular body and the glenoid. -

Evaluation of Humeral and Glenoid Bone Deformity in Glenohumeral Arthritis 5
Evaluation of Humeral and Glenoid Bone Deformity 1 in Glenohumeral Arthritis Brian F. Grogan and Charles M. Jobin Introduction glenoid bone wear helps the surgeon formulate a successful treatment plan and surgical goals Glenohumeral arthritis is the sequela of a vari- to address the pathoanatomy and improve the ety of pathologic shoulder processes, most durability of shoulder arthroplasty. The evalu- commonly degenerative osteoarthritis, but may ation of humeral and glenoid bone deformity also be secondary to post-traumatic conditions, in glenohumeral arthritis has profound surgical inflammatory arthritis, rotator cuff tear arthrop- implications and is fundamental to successful athy, and postsurgical conditions most com- shoulder arthroplasty. monly post-capsulorrhaphy arthritis. Patients with glenohumeral arthritis commonly demon- strate patterns of bony deformity on the glenoid Glenoid Deformity in Osteoarthritis and humerus that are caused by the etiology of the arthritis. For example, osteoarthritis com- Glenoid deformity and glenohumeral subluxation monly presents with posterior glenoid wear, are commonly seen in the setting of primary osteo- secondary glenoid retroversion, and posterior arthritis of the glenohumeral joint. The glenoid humeral head subluxation, while inflammatory wear tends to occur posteriorly and may be best arthritis routinely causes concentric glenoid viewed on axial radiographs or computed tomog- wear with central glenoid erosion. A thorough raphy (CT) axial images. Glenoid erosion, as first history and physical, as well as laboratory and characterized by Walch, is noted to be either central radiographic workup, are keys to understanding or posterior, with varying degrees of wear and pos- the etiology of arthritis and understanding the terior subluxation of the humerus [1, 2] (Fig. -

Integrating the Shoulder Complex to the Body As a Whole: Practical Applications for the Dancer
RESOURCE PAPER FOR TEACHERS INTEGRATING THE SHOULDER COMPLEX TO THE BODY AS A WHOLE: PRACTICAL APPLICATIONS FOR THE DANCER LISA DONEGAN SHOAF, DPT, PHD AND JUDITH STEEL MA, CMA WITH THE IADMS DANCE EDUCATORS’ COMMITTEE, 2018. TABLE OF CONTENTS 1. Introduction 2 2. Anatomy and Movements of the Shoulder Complex 3 3. Force Couples: Muscle Connections of the Scapula and Upper Extremity 12 4. The Shoulder Joint and Rotator Cuff Muscles 17 5. Integration of the Shoulder Girdle to the Trunk, Pelvis and Lower Extremities 20 6. Common Dancer Issues in the Upper Extremity 22 7. Summary 25 8. Illustration Credits 26 9. Recommended Reading 27 10. The Authors 28 1. INTRODUCTION Figure 1: Ease and elegance in the shoulder complex The focus in many forms of dance training is often on the movements of the lower body- the legs and feet. Movement of the upper body- trunk, shoulders, and arms, is often introduced later. Because so much emphasis is placed on function of the trunk and legs, and since most injuries for dancers occur in the lower body regions, it is easy to overlook the importance of optimal function of the shoulder and arms. The shoulder complex (defined as the humerus, clavicle, sternum, and scapula bones that form a girdle or shawl over the rib cage) in combination with its connection to the trunk and, ultimately, the lower body, has many components. There are three important concepts (listed below) that, when understood and experienced, can help dance educators and dancers better make the important connections to merge maximal range of motion with well aligned and supported movements that create full body artistic expressiveness. -

Scapular (Shoulder Blade) Disorders
DISEASES & CONDITIONS Scapular (Shoulder Blade) Disorders The scapula, or shoulder blade, is a large triangular-shaped bone that lies in the upper back. The bone is surrounded and supported by a complex system of muscles that work together to help you move your arm. If an injury or condition causes these muscles to become weak or imbalanced, it can alter the position of the scapula at rest or in motion. An alteration in scapular positioning or motion can make it difficult to move your arm, especially when performing overhead activities, and may cause your shoulder to feel weak. An alteration can also lead to injury if the normal ball-and-socket alignment of your shoulder joint is not maintained. Treatment for scapular disorders usually involves physical therapy designed to strengthen the muscles in the shoulder and restore the proper position and motion of the scapula. Anatomy Your shoulder joint is a ball-and-socket joint. The head of the humerus (upper arm bone) is the ball and the scapula (shoulder blade) forms the socket where the humerus sits. The scapula and arm are connected to the body by multiple muscle and ligament attachments. The front of the scapula (acromion) is also connected to the clavicle (collarbone) through the acromioclavicular joint. As you move your arm around your body, your scapula must also move to maintain the ball and socket in normal alignment. (Left) The bones of the shoulder. The scapula serves as a site for the attachment of multiple muscles around the shoulder. (Right) The muscles and soft tissues of the shoulder. -

Chapter 5 the Shoulder Joint
The Shoulder Joint • Shoulder joint is attached to axial skeleton via the clavicle at SC joint • Scapula movement usually occurs with movement of humerus Chapter 5 – Humeral flexion & abduction require scapula The Shoulder Joint elevation, rotation upward, & abduction – Humeral adduction & extension results in scapula depression, rotation downward, & adduction Manual of Structural Kinesiology – Scapula abduction occurs with humeral internal R.T. Floyd, EdD, ATC, CSCS rotation & horizontal adduction – Scapula adduction occurs with humeral external rotation & horizontal abduction © McGraw-Hill Higher Education. All rights reserved. 5-1 © McGraw-Hill Higher Education. All rights reserved. 5-2 The Shoulder Joint Bones • Wide range of motion of the shoulder joint in • Scapula, clavicle, & humerus serve as many different planes requires a significant attachments for shoulder joint muscles amount of laxity – Scapular landmarks • Common to have instability problems • supraspinatus fossa – Rotator cuff impingement • infraspinatus fossa – Subluxations & dislocations • subscapular fossa • spine of the scapula • The price of mobility is reduced stability • glenoid cavity • The more mobile a joint is, the less stable it • coracoid process is & the more stable it is, the less mobile • acromion process • inferior angle © McGraw-Hill Higher Education. All rights reserved. 5-3 © McGraw-Hill Higher Education. All rights reserved. From Seeley RR, Stephens TD, Tate P: Anatomy and physiology , ed 7, 5-4 New York, 2006, McGraw-Hill Bones Bones • Scapula, clavicle, & humerus serve as • Key bony landmarks attachments for shoulder joint muscles – Acromion process – Humeral landmarks – Glenoid fossa • Head – Lateral border • Greater tubercle – Inferior angle • Lesser tubercle – Medial border • Intertubercular groove • Deltoid tuberosity – Superior angle – Spine of the scapula © McGraw-Hill Higher Education. -
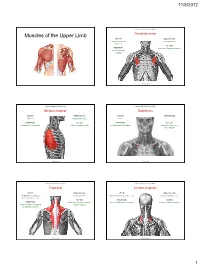
Muscles of the Upper Limb.Pdf
11/8/2012 Muscles Stabilizing Pectoral Girdle Muscles of the Upper Limb Pectoralis minor ORIGIN: INNERVATION: anterior surface of pectoral nerves ribs 3 – 5 ACTION: INSERTION: protracts / depresses scapula coracoid process (scapula) (Anterior view) Muscles Stabilizing Pectoral Girdle Muscles Stabilizing Pectoral Girdle Serratus anterior Subclavius ORIGIN: INNERVATION: ORIGIN: INNERVATION: ribs 1 - 8 long thoracic nerve rib 1 ---------------- INSERTION: ACTION: INSERTION: ACTION: medial border of scapula rotates scapula laterally inferior surface of scapula stabilizes / depresses pectoral girdle (Lateral view) (anterior view) Muscles Stabilizing Pectoral Girdle Muscles Stabilizing Pectoral Girdle Trapezius Levator scapulae ORIGIN: INNERVATION: ORIGIN: INNERVATION: occipital bone / spinous accessory nerve transverse processes of C1 – C4 dorsal scapular nerve processes of C7 – T12 ACTION: INSERTION: ACTION: INSERTION: stabilizes / elevates / retracts / upper medial border of scapula elevates / adducts scapula acromion / spine of scapula; rotates scapula lateral third of clavicle (Posterior view) (Posterior view) 1 11/8/2012 Muscles Stabilizing Pectoral Girdle Muscles Moving Arm Rhomboids Pectoralis major (major / minor) ORIGIN: INNERVATION: ORIGIN: INNERVATION: spinous processes of C7 – T5 dorsal scapular nerve sternum / clavicle / ribs 1 – 6 dorsal scapular nerve INSERTION: ACTION: INSERTION: ACTION: medial border of scapula adducts / rotates scapula intertubucular sulcus / greater tubercle flexes / medially rotates / (humerus) adducts