Appendix O20193
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Complete Issue
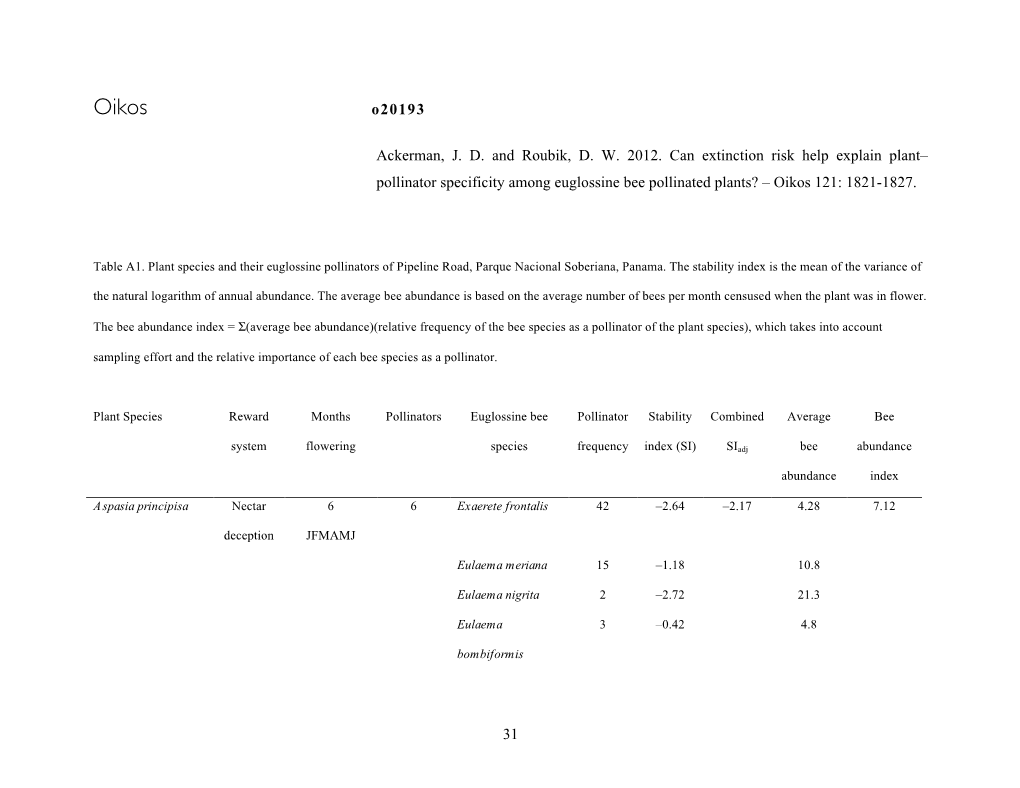
ISSN 1409-3871 VOL. 8, No. 2 AUGUST 2008 Capsule development, in vitro germination and plantlet acclimatization in Phragmipedium humboldtii, P. longifolium and P. pearcei MELANIA MUÑOZ & VÍCTOR M. JI M ÉNEZ 23 Stanhopeinae Mesoamericanae IV: las Coryanthes de Charles W. Powell GÜNTER GERLACH & GUSTA V O A. RO M ERO -GONZÁLEZ 33 The Botanical Cabinet RUDOLF JENNY 43 New species and records of Orchidaceae from Costa Rica DIE G O BO G ARÍN , ADA M KARRE M ANS & FRANCO PU P ULIN 53 Book reviews 75 THE INTERNATIONAL JOURNAL ON ORCHIDOLOGY LANKESTERIANA THE IN T ERNA ti ONAL JOURNAL ON ORCH I DOLOGY Copyright © 2008 Lankester Botanical Garden, University of Costa Rica Effective publication date: August 29, 2008 Layout: Jardín Botánico Lankester. Cover: Plant of Epidendrum zunigae Hágsater, Karremans & Bogarín. Drawing by D. Bogarín. Printer: Litografía Ediciones Sanabria S.A. Printed copies: 500 Printed in Costa Rica / Impreso en Costa Rica R Lankesteriana / The International Journal on Orchidology No. 1 (2001)-- . -- San José, Costa Rica: Editorial Universidad de Costa Rica, 2001-- v. ISSN-1409-3871 1. Botánica - Publicaciones periódicas, 2. Publicaciones periódicas costarricenses LANKESTERIANA 8(2): 23-31. 2008. CAPSULE DEVELOPMENT, IN VITRO GERMINATION AND PLANTLET ACCLIMATIZATION IN PHRAGMIPEDIUM HUMBOLDTII, P. LONGIFOLIUM AND P. PEARCEI MELANIA MUÑOZ 1 & VÍCTOR M. JI M ÉNEZ 2 CIGRAS, Universidad de Costa Rica, 2060 San Pedro, Costa Rica Jardín Botánico Lankester, Universidad de Costa Rica, P.O. Box 1031, 7050 Cartago, Costa Rica [email protected]; [email protected] ABSTRACT . Capsule development from pollination to full ripeness was evaluated in Phragmipedium longifolium, P. -

Santanderella, a Colombian New Genus in the Oncidiinae (Orchidaceae)
SANTANDERELLA, A COLOMBIAN NEW GENUS IN THE ONCIDIINAE (ORCHIDACEAE) Pedro Ortiz v. * Abstract: Santanderella, a new genus of orchids from Colombia, in the subtribe oncidiinae is proposed, with the type species, Santan- derella amado-rinconiana. Key words: Orchidaceae, Oncidiinae, new genus Santanderella, new species Santanderella amado-rinconiana, Colombia, plant taxo- nomy. For the Orchid show held by the Asociación Bogotana de Orquideología on [uly 2-5,2009, Mr Orlando Rincón, an orchid grower from Bucaramanga, Santander, brought a small orchid plant from Santander (Colombia), collected by the young Jonathan Amado, in Floridablanca, near Bucaramanga (Santander, Colombia). This plant had been collected previously by Orlando Rincón in the same area. The plant cau~ht our attention, not because of its size or the extraordinary beauty of its flowers" but due to its peculiar characters. Unfortunately during the days of the show the flowers did not open, and so it was not possible to come to a sure identification. Orlando left us the little plant so that we might be able to study it at ease, a task I immediately undertook. With the aid of friends, good photo- graphers, we managed to assemble a good documentation of the characteristic features of this orchid. Above all, it was clear that it belongs to the subtribe On- cidiinae (sensu R. Dressler, 1981) and among this large subtribe it shows affinity with the genera Notylia and Macroclinium. The genera of this group have been defined and characterized in different ways, as can be seen in the study publis- hed by F. Pupulin (1997)1, to which we refer for further information. -

Los Tuxtlas", Veracruz, México: (Violaceaea-Zingiberaceae)
Rev. Biol. Trop., 44(2): 427-447, 1996 Lista comentada de plantas de la Estación de Biología Tropical "Los Tuxtlas", Veracruz, México: (Violaceaea-Zingiberaceae) Guillermo Ibarra-Manríquez y Santiago Sinaca Colín Departamento de Botánica, Instituto de Biología, Universidad Nacional Autónoma de México, Apartado Postal 70-233. México 045 10. D. F. Abstrad: This is the third and final part of the commented list of plants found in Los Tuxtlas, a Mexican Tropical Biology Research Station.Previous parts appeared in Revista de Biologia Tropical vol. 43 (1-3) and vol. 44 (1). Key words; Phenology, vascular plants, floristics, Tropical rain forest, uses, Mexico, Veracruz. Esta es la tercera y última parte de la lista pálido, lustroso, pecíolo fr ecuent emente florística de la Estación de Biología Tropical recurvado, rojo en hojas jóvenes y verde gris "Los Tuxtlas"; las 663 primeras partes en la hoj a madura, sin domacios, costa aparecieron en Revista de Biología Tropical vol. glabra; flor blanca, cápsula verde, con la 43 (1-3) y 44 (1). semilla subglobosa, pelosa. FI: febrero a mayo. Fr : marzo a septiembre. ALG 32534, VIOLACEAE 32596, GHN 20351, GIM 80, 459, 517, 991, 1322, 1562, 1578, 1979, 2337, ICA 693, * Hybanthus attenuatus (Kunth) Schulze MRR 1225, 1387, MSS 11966, RCT 2660, Menz Herbácea de hasta 0.5 m; florblanca, con WEB 20917. la gargantade color lila, cápsula verde, Fl y Fr: enero a diciembre. GIM3428 , SSC 769, 1962. VITACEAE Orthion oblanceolatum Lundell Arbol de 6- 10 (-15) m, hojas simples, alternas, margen con Cissus gossypifolia Standl. Liana, hojas sim dientes inconspicuos, a manera de puntos ples, alternas; flor roja, baya negra. -

Capítulo 4. Biodiversidad
Biodiversidad El cambio climático global y la pérdida de la biodiversidad son dos de los problemas ambientales más importantes que enfrenta la humanidad hoy día. La expansión e intensificación de las actividades humanas desde mediados del siglo pasado han cambiado radicalmente el funcionamiento en muchos ecosistemas en diversas regiones del mundo e, incluso, han alterado los patrones de biodiversidad a nivel local y regional. En algunos casos, estos cambios han provocado la extinción de muchas especies. Estimaciones sugieren que la tasa actual de extinción de especies a nivel mundial podría ser entre 10 y 1 000 veces mayor a la registrada con anterioridad a la presencia humana (Pimm et al., 1995). No obstante, los grandes esfuerzos que se han hecho en las últimas dos décadas a nivel internacional para conservar y utilizar sustentablemente la biodiversidad, han sido insuficientes para avanzar hacia el cumplimiento de las Metas de Aichi para la Diversidad Biológica (SCBD, 2014). De no haber cambios importantes, los pronósticos basados en la proyección de las tendencias actuales de los factores de presión más importantes, sugieren que la pérdida de la riqueza biológica global podría continuar cuando menos hasta el año 2020 (SCDB, 2010). Los efectos de la pérdida de la biodiversidad no se restringen al aspecto ambiental. Es ampliamente reconocido que el bienestar social y el desarrollo económico de las naciones, y en particular el de los países en desarrollo y el de las comunidades más vulnerables, están fincados en la continuidad de los servicios ambientales que brindan los ecosistemas y su biodiversidad (MEA, 2005). Para muchos países, incluido México, la pérdida de biodiversidad cobra mayor relevancia debido a que es considerado como uno de los centros de diversidad biológica más importantes del planeta, de hecho, forma parte del grupo de los 15 países megadiversos que concentran en conjunto entre el 60 y 70% de la biodiversidad global (Mittermeier et al., 1997; UNEP 2010). -

Pollinator Specificity and Seasonal Patterns in the Euglossine Bee-Orchid Mutualism at La Gamba Biological Station
Acta ZooBot Austria 156, 2019, 171–181 Pollinator specificity and seasonal patterns in the euglossine bee-orchid mutualism at La Gamba Biological Station Santiago R. Ramirez The plant family Orchidaceae exhibits some of the most spectacular and intricate ad- aptations for insect pollination. Across the Neotropical region male euglossine bees provide pollination services to approx. 700 orchid species that have evolved scent pro- duction in exchange for sexual reproduction. Male orchid bees collect scents from flowers and other sources to concoct perfume mixtures that they use as pheromone analogs during courtship display. Although the pollination biology of some of these associations has been studied in detail for some orchid taxa, community-wide analyses of this mutualism are lacking. Here I present an analysis of the plant-pollinator affilia- tion patterns and phenology among scent-producing orchids and male euglossine bees based on 960 bee-orchid interactions obtained over the course of five years of sampling at La Gamba Biological Station (south-western Costa Rica). I identify a highly nested plant-pollinator network that is composed of 24 bee species and 17 orchid genera. Some orchid genera exhibit pronounced flowering seasonality, with most of the diver- sity of interactions taking place during the dry season (March-April) and few orchid taxa blooming throughout the year. The architecture of the plant-pollinator network also revealed a substantial degree of pollinator sharing among orchid genera, suggest- ing that distantly related lineages independently converged on the use of similar pol- linator bee assemblages. RAMIREZ S.R., 2019: Bestäuberspezifizität und jahreszeitliche Variation in Pracht- bienen-Orchideen Mutualismen an der Tropenstation La Gamba. -

The Orchid Flora of the Colombian Department of Valle Del Cauca Revista Mexicana De Biodiversidad, Vol
Revista Mexicana de Biodiversidad ISSN: 1870-3453 [email protected] Universidad Nacional Autónoma de México México Kolanowska, Marta The orchid flora of the Colombian Department of Valle del Cauca Revista Mexicana de Biodiversidad, vol. 85, núm. 2, 2014, pp. 445-462 Universidad Nacional Autónoma de México Distrito Federal, México Available in: http://www.redalyc.org/articulo.oa?id=42531364003 How to cite Complete issue Scientific Information System More information about this article Network of Scientific Journals from Latin America, the Caribbean, Spain and Portugal Journal's homepage in redalyc.org Non-profit academic project, developed under the open access initiative Revista Mexicana de Biodiversidad 85: 445-462, 2014 Revista Mexicana de Biodiversidad 85: 445-462, 2014 DOI: 10.7550/rmb.32511 DOI: 10.7550/rmb.32511445 The orchid flora of the Colombian Department of Valle del Cauca La orquideoflora del departamento colombiano de Valle del Cauca Marta Kolanowska Department of Plant Taxonomy and Nature Conservation, University of Gdańsk. Wita Stwosza 59, 80-308 Gdańsk, Poland. [email protected] Abstract. The floristic, geographical and ecological analysis of the orchid flora of the department of Valle del Cauca are presented. The study area is located in the southwestern Colombia and it covers about 22 140 km2 of land across 4 physiographic units. All analysis are based on the fieldwork and on the revision of the herbarium material. A list of 572 orchid species occurring in the department of Valle del Cauca is presented. Two species, Arundina graminifolia and Vanilla planifolia, are non-native elements of the studied orchid flora. The greatest species diversity is observed in the montane regions of the study area, especially in wet montane forest. -

Additions to the Orchid Flora of Belize, Central America
LANKESTERIANA 8: 1-3. 2003. ADDITIONS TO THE ORCHID FLORA OF BELIZE, CENTRAL AMERICA BRENDAN SAYERS1 and HEATHER DU PLOOY2 1 National Botanic Gardens, Glasnevin, Dublin 9, Ireland 2 Belize Botanic Gardens, San Ignacio, Cayo, Belize AB S T R A C T. The National Botanic Gardens, Glasnevin, Dublin, Ireland have travelled to Belize, Central America on five expeditions for the main purpose of collecting living specimens of Orchidaceae. Four of the expeditions were in conjunction with Belize Botanic Gardens. As a result of these expeditions 17 new records for Orchidaceae are being added to the published flora. RE S U M E N. Los Jardines Botánicos Nacionales de Glasnevin, Dublín, Irlanda, han realizado cinco expedi- ciones a Belice, América Central, con el objetivo principal de recolectar especímenes vivos de Orchidaceae. Cuatro de las expediciones se realizaron en conjunto con el Jardín Botanico de Belice. Cómo resultado de estas expediciones se añadieron a las floras publicadas 17 nuevos registros de Orchidaceae. PALABRAS CLAVE / KEY WORDS: Orchidaceae, Belice, Nuevos registros. I n t r o d u c t i o n. The National Botanic Gardens, cochleata (L.) Lemée var. cochleata. Glasnevin (NBGG) expeditions took place on an To date 17 of the specimens have been identified annual basis from 1996 to 2000. Brendan as new records of Orchidaceae for the Belizean flora. O'Donoghue, a past student of the NBBG, assisted the Some of these records have been included in reports 1996 expedition, the 1997 expedition was assisted by of the expeditions lodged in the Library, National Brendan O’Donoghue and Belize Botanic Gardens Botanic Gardens, Glasnevin, Ireland and with the and the 1998, 1999 and 2000 were joint expeditions Department of Forestry and Natural Resources, with Belize Botanic Gardens. -

The Genus Systeloglossum
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/256404679 The Genus Systeloglossum Article · January 1970 CITATION READS 1 60 2 authors: Robert Dressler Norris H Williams University of Costa Rica University of Florida 355 PUBLICATIONS 6,235 CITATIONS 131 PUBLICATIONS 4,002 CITATIONS SEE PROFILE SEE PROFILE Some of the authors of this publication are also working on these related projects: Sobralias of Costa Rica View project Determination of the new species of Sobralia found growing in the Turrialba Valley View project All content following this page was uploaded by Robert Dressler on 27 May 2014. The user has requested enhancement of the downloaded file. The Genus Systeloglossum ROBERT L. DRESSLER AND NORRIS H. WILLIAMS HE COMPLEX OF GENERA which make up the subtribe Oncidiinae are notable for their great diversity of flower structure. It is not surprising T that Schlechter grouped these genera into as many as ten different btribes. How can one imagine close relationships between Notylia and ~richoPilia, or Quekettia and Odontoglossum? In spite of their striking differences, we now group them all into the subtribe Oncidiinae, for two 'mportant reasons. First, crossing experiments, especially those of Mr. Moir, ~ave shown that the whole group is closely tied together by interfertility. We may not be able to cross Comparettia with Brassia, but we can cross both with Oncidium. The second reason for grouping them together is that even the morphological differences do not hold up very well on close inspection. Notylia may seem very different from Trichopilia, but Notylia and Macradenia are surely closely related, and Macradenia and Trichopilia subulata are perhaps more similar to each other than T . -

Phylogeny, Genome Size, and Chromosome Evolution of Asparagales J
Aliso: A Journal of Systematic and Evolutionary Botany Volume 22 | Issue 1 Article 24 2006 Phylogeny, Genome Size, and Chromosome Evolution of Asparagales J. Chris Pires University of Wisconsin-Madison; University of Missouri Ivan J. Maureira University of Wisconsin-Madison Thomas J. Givnish University of Wisconsin-Madison Kenneth J. Systma University of Wisconsin-Madison Ole Seberg University of Copenhagen; Natural History Musem of Denmark See next page for additional authors Follow this and additional works at: http://scholarship.claremont.edu/aliso Part of the Botany Commons Recommended Citation Pires, J. Chris; Maureira, Ivan J.; Givnish, Thomas J.; Systma, Kenneth J.; Seberg, Ole; Peterson, Gitte; Davis, Jerrold I.; Stevenson, Dennis W.; Rudall, Paula J.; Fay, Michael F.; and Chase, Mark W. (2006) "Phylogeny, Genome Size, and Chromosome Evolution of Asparagales," Aliso: A Journal of Systematic and Evolutionary Botany: Vol. 22: Iss. 1, Article 24. Available at: http://scholarship.claremont.edu/aliso/vol22/iss1/24 Phylogeny, Genome Size, and Chromosome Evolution of Asparagales Authors J. Chris Pires, Ivan J. Maureira, Thomas J. Givnish, Kenneth J. Systma, Ole Seberg, Gitte Peterson, Jerrold I. Davis, Dennis W. Stevenson, Paula J. Rudall, Michael F. Fay, and Mark W. Chase This article is available in Aliso: A Journal of Systematic and Evolutionary Botany: http://scholarship.claremont.edu/aliso/vol22/iss1/ 24 Asparagales ~£~2COTSgy and Evolution Excluding Poales Aliso 22, pp. 287-304 © 2006, Rancho Santa Ana Botanic Garden PHYLOGENY, GENOME SIZE, AND CHROMOSOME EVOLUTION OF ASPARAGALES 1 7 8 1 3 9 J. CHRIS PIRES, • • IVAN J. MAUREIRA, THOMAS J. GIVNISH, 2 KENNETH J. SYTSMA, 2 OLE SEBERG, · 9 4 6 GITTE PETERSEN, 3· JERROLD I DAVIS, DENNIS W. -

Common Epiphytes and Lithophytes of BELIZE 1 Bruce K
Common Epiphytes and Lithophytes of BELIZE 1 Bruce K. Holst, Sally Chambers, Elizabeth Gandy & Marilynn Shelley1 David Amaya, Ella Baron, Marvin Paredes, Pascual Garcia & Sayuri Tzul2 1Marie Selby Botanical Gardens, 2 Ian Anderson’s Caves Branch Botanical Garden © Marie Selby Bot. Gard. ([email protected]), Ian Anderson’s Caves Branch Bot. Gard. ([email protected]). Photos by David Amaya (DA), Ella Baron (EB), Sally Chambers (SC), Wade Coller (WC), Pascual Garcia (PG), Elizabeth Gandy (EG), Bruce Holst (BH), Elma Kay (EK), Elizabeth Mallory (EM), Jan Meerman (JM), Marvin Paredes (MP), Dan Perales (DP), Phil Nelson (PN), David Troxell (DT) Support from the Marie Selby Botanical Gardens, Ian Anderson’s Caves Branch Jungle Lodge, and many more listed in the Acknowledgments [fieldguides.fieldmuseum.org] [1179] version 1 11/2019 TABLE OF CONTENTS long the eastern slopes of the Andes and in Brazil’s Atlantic P. 1 ............. Epiphyte Overview Forest biome. In these places where conditions are favorable, epiphytes account for up to half of the total vascular plant P. 2 .............. Epiphyte Adaptive Strategies species. Worldwide, epiphytes account for nearly 10 percent P. 3 ............. Overview of major epiphytic plant families of all vascular plant species. Epiphytism (the ability to grow P. 6 .............. Lesser known epiphytic plant families as an epiphyte) has arisen many times in the plant kingdom P. 7 ............. Common epiphytic plant families and species around the world. (Pteridophytes, p. 7; Araceae, p. 9; Bromeliaceae, p. In Belize, epiphytes are represented by 34 vascular plant 11; Cactaceae, p. 15; p. Gesneriaceae, p. 17; Orchida- families which grow abundantly in many shrublands and for- ceae, p. -

Comparative Vegetative Anatomy and Systematics of Oncidiinae (Maxillarieae, Orchidaceae) William Louis Stern University of Florida
CORE Metadata, citation and similar papers at core.ac.uk Provided by The Keep Eastern Illinois University The Keep Faculty Research & Creative Activity Biological Sciences January 2006 Comparative vegetative anatomy and systematics of Oncidiinae (Maxillarieae, Orchidaceae) William Louis Stern University of Florida Barbara S. Carlsward Eastern Illinois University, [email protected] Follow this and additional works at: http://thekeep.eiu.edu/bio_fac Part of the Biology Commons Recommended Citation Stern, William Louis and Carlsward, Barbara S., "Comparative vegetative anatomy and systematics of Oncidiinae (Maxillarieae, Orchidaceae)" (2006). Faculty Research & Creative Activity. 263. http://thekeep.eiu.edu/bio_fac/263 This Article is brought to you for free and open access by the Biological Sciences at The Keep. It has been accepted for inclusion in Faculty Research & Creative Activity by an authorized administrator of The Keep. For more information, please contact [email protected]. Comparative vegetative anatomy and systematics of the Oncidiinae (Maxillarieae, Orchidaceae) WILLIAM LOUIS STERN and BARBARA S. CARLSWARD Abstract Subtribe Oncidiinae comprises a vegetatively heterogeneous assemblage of species that has persistently been incapable of organization. Anatomy was considered to be a possible means to resolve the perplexity of relationships amongst the constituent taxa. The consistent occurrence of a foliar hypodermis, homogeneous mesophyll, conical silica bodies in stegmata, and ubiquitous fibre bundles in leaves provides a matrix for linKing the taxa, as do the parenchymatous pith and O-thickened endodermal cell walls in roots. However, the strict consensus of the 40 genera studied was completely unresolved, suggesting that vegetative characters alone are insufficient to assess the relationships amongst these taxa, a conclusion also reached for the remainder of Maxillarieae. -

Expansion of the Geographical Distribution of Notylia Angustifolia (Orchidaceae): First Record for the Amazonas State (AM), Brazila
Expansion of the geographical distribution of Notylia angustifolia (Orchidaceae): first record for the Amazonas state (AM), Brazila Amauri Herbert Krahl1,*, Jefferson José Valsko2, Dayse Raiane Passos Krahl3, Ana Sofia Sousa de Holanda1 & Renato Ximenes Bolsanello4,* Abstract Notylia angustifolia Cogniaux is reported for the first time for the Brazilian State of Amazonas. Résumé Notylia angustifolia (Orchidaceae : premier enregistrement pour l'état d'Amazonas (Brésil) – Notylia angustifolia Cogniaux, connu au Brésil dans les états de Roraima et Rondania, est ici rapporté pour l'état d'Amazonas. The Neotropical genus Notylia, proposed by Lindley in 1825, is comprised of about 60 species (Chase et al., 2003), of which 25 can be found in Brazil, 15 in the Brazilian Amazon and 7 in the State of Amazonas (Barros et al., 2015). Notylia angustifolia Cogniaux has recently been reported from Brazil by Pessoa et al. (2013), more precisely from the Parque Nacional do Viruá, in the State of Roraima (RR). This species also occurs in the State of Rondônia (RO), near the Rio Jarú, município Jí-paraná (Barros et al., 2015). The goal of the present note is to report the first record of the species for the State of Amazonas (AM - Fig. 1). Material: BRASIL; Amazonas: Manaus, BR 174, KM 44, Reserva Biológica da Campina, Campina aberta, 10.IX.2011, floresceu em cultivo em 16. XII.2014, A. H. Krahl & J. J. Valsko 486 (EAFM); Novo Airão, KM 27, campinarana, 13.XII.2014, A. H. Krahl & J. J. Valsko 485 (INPA). a : manuscrit reçu le 22 juillet 2015, accepté le 11 août 2015 article mis en ligne sur www.richardiana.com le 12/08/2015 – pp.