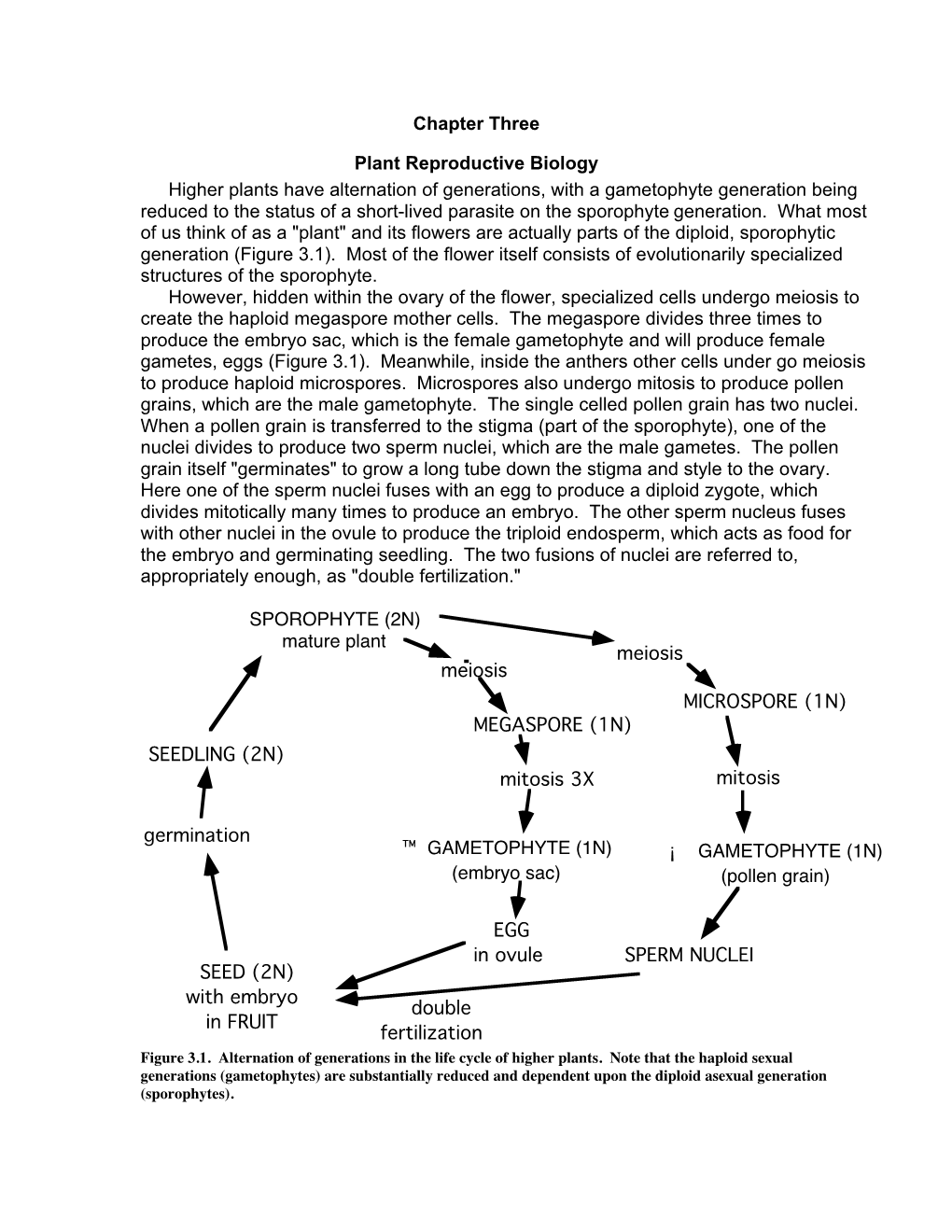
Chapter Three Plant Reproductive Biology Higher Plants Have Alternation of Generations, with a Gametophyte Generation Being Redu
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Inflorescences
Inflorescences - Floral Displays Inflorescences - Floral Displays A shift from widely The vast majority of flowering plants spaced single flowers to an inflorescence required possess flowers in clusters called an condensation of shoots inflorescence. and the loss of the intervening leaves. These clusters facilitate pollination via a prominent visual display and more The simplest efficient pollen uptake and deposition. inflorescence type would thus be indeterminate with the oldest flowers at the base and the younger flowers progressively closer to the apical meristem of the shoot. = a raceme Raceme (Prunus or cherry) One modification of the basic raceme is to make it compound The panicle is essentially compound a series of attached racemes with the oldest racemes at the base and the youngest at the apex of the inflorescence. Panicle (Zigadenus or white camass) Raceme Panicle 1 A second modification of the basic raceme is to lose its pedicels The spike is usually associated with congested reduced flowers and often, but not always, with wind Pedicel loss pollination. wind pollinated Spike (Plantago or plantain) Raceme Spike A third modification of the basic raceme is to lose its internodes The spike is usually associated with congested reduced flowers and often, animal pollinated but not always, with wind Internode loss pollination. Umbel Spike (Combretum - Brent’s plants) Raceme 2 The umbel characterizes specific families (carrot and The umbel is found scattered in many other families ginseng families for example). as well. These families typically show a compound umbel - smaller umbellets on a larger umbel. Umbel Umbel (Cicuta or water hemlock) (Zizia or golden alexander) (Eriogonum or false buckwheat - family Polygonaceae) - Ben’s plants A fourth modification of the basic raceme is for the stem axis to form a head The head or capitulum characterizes specific families - most notably the Compositae or Asteraceae. -

1 5 Chapter 1 Introduction I. LIFE CYCLES and DIVERSITY of VASCULAR PLANTS the Subjects of This Thesis Are the Pteri
Chapter 1-15 Chapter 1 Introduction I. LIFE CYCLES AND DIVERSITY OF VASCULAR PLANTS The subjects of this thesis are the pteridophytes and seed plants that are conventionally classified as the vascular plants or tracheophytes. Vascular plants were traditionally defined by the possession of specialized conducting tissues, called phloem and xylem. Mosses are now believed to have inherited their conducting tissues from a common ancestor with the tracheophytes (Mishler & Churchill 1984) but are not considered in this thesis. Vascular plants can be divided into four groups with respect to life cycle. These groups are homosporous pteridophytes, heterosporous pteridophytes, gymnosperms and angiosperms. This is not intended to be a phylogenetic classification. There are about a quarter of a million species of vascular plant alive today. The vast majority are angiosperms and most of the remainder are homosporous pteridophytes. Heterosporous pteridophytes and gymnosperms contribute only a small number of species (Table 1.1). TABLE 1.1 Estimated number of extant species in each of the major groups of vascular plants (data from Parker 1982). Homosporous pteridophytes 12,000 species Heterosporous pteridophytes <1,000 species Gymnosperms <1,000 species Angiosperms >200,000 species A. Homosporous pteridophytes Homosporous pteridophytes produce a single type of spore. Spores are dispersed and develop into photosynthetic or mycoparasitic gametophytes. A gametophyte's gender is indeterminate at the time of spore dispersal, and a single gametophyte may produce eggs and/or sperm. Sperm are motile, and require free water to fertilize eggs. The young sporophyte is nourished by the maternal gametophyte during early development but later becomes Chapter 1-16 nutritionally independent. -

Gymnosperms the MESOZOIC: ERA of GYMNOSPERM DOMINANCE
Chapter 24 Gymnosperms THE MESOZOIC: ERA OF GYMNOSPERM DOMINANCE THE VASCULAR SYSTEM OF GYMNOSPERMS CYCADS GINKGO CONIFERS Pinaceae Include the Pines, Firs, and Spruces Cupressaceae Include the Junipers, Cypresses, and Redwoods Taxaceae Include the Yews, but Plum Yews Belong to Cephalotaxaceae Podocarpaceae and Araucariaceae Are Largely Southern Hemisphere Conifers THE LIFE CYCLE OF PINUS, A REPRESENTATIVE GYMNOSPERM Pollen and Ovules Are Produced in Different Kinds of Structures Pollination Replaces the Need for Free Water Fertilization Leads to Seed Formation GNETOPHYTES GYMNOSPERMS: SEEDS, POLLEN, AND WOOD THE ECOLOGICAL AND ECONOMIC IMPORTANCE OF GYMNOSPERMS The Origin of Seeds, Pollen, and Wood Seeds and Pollen Are Key Reproductive SUMMARY Innovations for Life on Land Seed Plants Have Distinctive Vegetative PLANTS, PEOPLE, AND THE Features ENVIRONMENT: The California Coast Relationships among Gymnosperms Redwood Forest 1 KEY CONCEPTS 1. The evolution of seeds, pollen, and wood freed plants from the need for water during reproduction, allowed for more effective dispersal of sperm, increased parental investment in the next generation and allowed for greater size and strength. 2. Seed plants originated in the Devonian period from a group called the progymnosperms, which possessed wood and heterospory, but reproduced by releasing spores. Currently, five lineages of seed plants survive--the flowering plants plus four groups of gymnosperms: cycads, Ginkgo, conifers, and gnetophytes. Conifers are the best known and most economically important group, including pines, firs, spruces, hemlocks, redwoods, cedars, cypress, yews, and several Southern Hemisphere genera. 3. The pine life cycle is heterosporous. Pollen strobili are small and seasonal. Each sporophyll has two microsporangia, in which microspores are formed and divide into immature male gametophytes while still retained in the microsporangia. -

BIL 161: Environment and Development: the Effects of Environmental Variables on Seed Germination
BIL 161: Environment and Development: The Effects of Environmental Variables on Seed Germination The seed is more than just a plant waiting to happen. It is a complex marvel of evolution, a miniature life-support system that responds to environmental cues in order to give the embryo nestled within the best chance of survival. I. Characteristics and Classification of Plants Plants share synapomorphies that set them apart from other organisms. 1. true tissues (of types unique to plants) 2. waxy cuticle (to prevent desiccation) 3. stomates (microscopic gas exchange pores on the leaves) 4. apical meristems (permanent embryonic tissue for constant growth) 5. multicellular sex organs (male antheridia and female archegonia) 6. walled spores produced in structures called sporangia 7. embryo development inside the female parent 8. secondary metabolites (alkaloids, tannins, flavonoids, etc.) 9. heteromorphic alternation of generations The most primitive plants do not produce seeds at all, but rather release spores into the environment where they grow into a second life cycle stage, called the gametophyte. In seed plants, the life cycle is highly derived. Seed plants still make spores, but each spore grows into a gametophyte that is little more than a bit of tissue that gives rise to gametes. In the male parts of the plant, each spore develops into a sperm-producing male gametophyte known as pollen. In the female parts of the plant, meiosis occurs inside a structure known as the ovule, which will eventually give rise to the seed. Plants can broadly be classified as follows. A. Bryophytes – non-vascular plants (mosses, liverworts and hornworts) B. -

Reproduction in Plants Which But, She Has Never Seen the Seeds We Shall Learn in This Chapter
Reproduction in 12 Plants o produce its kind is a reproduction, new plants are obtained characteristic of all living from seeds. Torganisms. You have already learnt this in Class VI. The production of new individuals from their parents is known as reproduction. But, how do Paheli thought that new plants reproduce? There are different plants always grow from seeds. modes of reproduction in plants which But, she has never seen the seeds we shall learn in this chapter. of sugarcane, potato and rose. She wants to know how these plants 12.1 MODES OF REPRODUCTION reproduce. In Class VI you learnt about different parts of a flowering plant. Try to list the various parts of a plant and write the Asexual reproduction functions of each. Most plants have In asexual reproduction new plants are roots, stems and leaves. These are called obtained without production of seeds. the vegetative parts of a plant. After a certain period of growth, most plants Vegetative propagation bear flowers. You may have seen the It is a type of asexual reproduction in mango trees flowering in spring. It is which new plants are produced from these flowers that give rise to juicy roots, stems, leaves and buds. Since mango fruit we enjoy in summer. We eat reproduction is through the vegetative the fruits and usually discard the seeds. parts of the plant, it is known as Seeds germinate and form new plants. vegetative propagation. So, what is the function of flowers in plants? Flowers perform the function of Activity 12.1 reproduction in plants. Flowers are the Cut a branch of rose or champa with a reproductive parts. -

Seed and Seed Dispersal
1st GRADE SEEDS AND SEED DISPERSAL Summary: This lab is all about seeds. First, students take apart a swollen lima bean seed and find the seed coat, food storage area, and the plant embryo. Second, the students sort a bag of seeds into groups and notice that all seeds look different but have the same three seed parts. Finally, students sort seeds that are dispersed in different ways. Students identify seeds that are dispersed by wind, hitchiking, animals carrying and burying, and animals eating and pooping. Intended Learning Outcomes for 1st Grade: Objective 1: Framing questions. Conducting investigations. Drawing conclusions. Objective 2: Developing social interaction skills with peers. Sharing ideas with peers. Connecting ideas with reasons. Objective 3: Ideas are supported by reasons. Communicaiton of ideas in science is important for helping to check the reasons for ideas. Utah State Core Curriculum Tie: Standard 4 Objective 1: Life Science Analyze the individual similarities and differences within and across larger groups. Standard 4 Objective 2: Life Science Describe and model life cycles of living things. Make observations about living things and their environment using the five senses. Preparation time: 1 hour to locate seeds the first time, then 20 min if seeds are reused Lesson time: 50 min Small group size: works best with one adult for every 5 students Materials: 1. one petri dish or paper towel per student 2. 1 bag of dried lima beans 3. One seed classification bag per group, this should include 5-6 seeds of about 15 different seed types. Use old seeds from seed packets or spices or seeds or nuts you may have in your kitchen. -

Auxin Regulation Involved in Gynoecium Morphogenesis of Papaya Flowers
Zhou et al. Horticulture Research (2019) 6:119 Horticulture Research https://doi.org/10.1038/s41438-019-0205-8 www.nature.com/hortres ARTICLE Open Access Auxin regulation involved in gynoecium morphogenesis of papaya flowers Ping Zhou 1,2,MahparaFatima3,XinyiMa1,JuanLiu1 and Ray Ming 1,4 Abstract The morphogenesis of gynoecium is crucial for propagation and productivity of fruit crops. For trioecious papaya (Carica papaya), highly differentiated morphology of gynoecium in flowers of different sex types is controlled by gene networks and influenced by environmental factors, but the regulatory mechanism in gynoecium morphogenesis is unclear. Gynodioecious and dioecious papaya varieties were used for analysis of differentially expressed genes followed by experiments using auxin and an auxin transporter inhibitor. We first compared differential gene expression in functional and rudimentary gynoecium at early stage of their development and detected significant difference in phytohormone modulating and transduction processes, particularly auxin. Enhanced auxin signal transduction in rudimentary gynoecium was observed. To determine the role auxin plays in the papaya gynoecium, auxin transport inhibitor (N-1-Naphthylphthalamic acid, NPA) and synthetic auxin analogs with different concentrations gradient were sprayed to the trunk apex of male and female plants of dioecious papaya. Weakening of auxin transport by 10 mg/L NPA treatment resulted in female fertility restoration in male flowers, while female flowers did not show changes. NPA treatment with higher concentration (30 and 50 mg/L) caused deformed flowers in both male and female plants. We hypothesize that the occurrence of rudimentary gynoecium patterning might associate with auxin homeostasis alteration. Proper auxin concentration and auxin homeostasis might be crucial for functional gynoecium morphogenesis in papaya flowers. -

Pollen Tube Guidance by Pistils Ensures Successful Double Fertilization Ravishankar Palanivelu∗ and Tatsuya Tsukamoto
Advanced Review Pathfinding in angiosperm reproduction: pollen tube guidance by pistils ensures successful double fertilization Ravishankar Palanivelu∗ and Tatsuya Tsukamoto Sexual reproduction in flowering plants is unique in multiple ways. Distinct multicellular gametophytes contain either a pair of immotile, haploid male gametes (sperm cells) or a pair of female gametes (haploid egg cell and homodiploid central cell). After pollination, the pollen tube, a cellular extension of the male gametophyte, transports both male gametes at its growing tip and delivers them to the female gametes to affect double fertilization. The pollen tube travels a long path and sustains its growth over a considerable amount of time in the female reproductive organ (pistil) before it reaches the ovule, which houses the female gametophyte. The pistil facilitates the pollen tube’s journey by providing multiple, stage-specific, nutritional, and guidance cues along its path. The pollen tube interacts with seven different pistil cell types prior to completing its journey. Consequently, the pollen tube has a dynamic gene expression program allowing it to continuously reset and be receptive to multiple pistil signals as it migrates through the pistil. Here, we review the studies, including several significant recent advances, that led to a better understanding of the multitude of cues generated by the pistil tissues to assist the pollen tube in delivering the sperm cells to the female gametophyte. We also highlight the outstanding questions, draw attention to opportunities created by recent advances and point to approaches that could be undertaken to unravel the molecular mechanisms underlying pollen tube–pistil interactions. 2011 Wiley Periodicals, Inc. -

Ap09 Biology Form B Q2
AP® BIOLOGY 2009 SCORING GUIDELINES (Form B) Question 2 Discuss the patterns of sexual reproduction in plants. Compare and contrast reproduction in nonvascular plants with that in flowering plants. Include the following topics in your discussion: (a) alternation of generations (b) mechanisms that bring female and male gametes together (c) mechanisms that disperse offspring to new locations Four points per part. Student must write about all three parts for full credit. Within each part it is possible to get points for comparing and contrasting. Also, specific points are available from details provided about nonvascular and flowering plants. Discuss the patterns of sexual reproduction in plants (4 points maximum): (a) Alternation of generations (4 points maximum): Topic Description (1 point each) Alternating generations Haploid stage and diploid stage. Gametophyte Haploid-producing gametes. Dominant in nonvascular plants. Double fertilization in flowering plants. Gametangia; archegonia and antheridia in nonvascular plants. Sporophyte Diploid-producing spores. Heterosporous in flowering plants. Flowering plants produce seeds; nonvascular plants do not. Flowering plants produce flower structures. Sporangia (megasporangia and microsporangia). Dominant in flowering plants. (b) Mechanisms that bring female and male gametes together (4 points maximum): Nonvascular Plants (1 point each) Flowering Plants (1 point each) Aquatic—requires water for motile sperm Terrestrial—pollination by wind, water, or animal Micropyle in ovule for pollen tube to enter Pollen tube to carry sperm nuclei Self- or cross-pollination Antheridia produce sperm Gametophytes; no antheridia or archegonia Archegonia produce egg Ovules produce female gametophytes/gametes Pollen: male gametophyte that produces gametes © 2009 The College Board. All rights reserved. Visit the College Board on the Web: www.collegeboard.com. -

Starch Biosynthesis in the Developing Endosperms of Grasses and Cereals
Review Starch Biosynthesis in the Developing Endosperms of Grasses and Cereals Ian J. Tetlow * and Michael J. Emes Department of Molecular and Cellular Biology, College of Biological Science, University of Guelph, Guelph, ON N1G 2W1, Canada; [email protected] * Correspondence: [email protected]; Tel.: +1-519-824-4120 Received: 31 October 2017; Accepted: 27 November 2017; Published: 1 December 2017 Abstract: The starch-rich endosperms of the Poaceae, which includes wild grasses and their domesticated descendents the cereals, have provided humankind and their livestock with the bulk of their daily calories since the dawn of civilization up to the present day. There are currently unprecedented pressures on global food supplies, largely resulting from population growth, loss of agricultural land that is linked to increased urbanization, and climate change. Since cereal yields essentially underpin world food and feed supply, it is critical that we understand the biological factors contributing to crop yields. In particular, it is important to understand the biochemical pathway that is involved in starch biosynthesis, since this pathway is the major yield determinant in the seeds of six out of the top seven crops grown worldwide. This review outlines the critical stages of growth and development of the endosperm tissue in the Poaceae, including discussion of carbon provision to the growing sink tissue. The main body of the review presents a current view of our understanding of storage starch biosynthesis, which occurs inside the amyloplasts of developing endosperms. Keywords: amylopectin; amylose; cereals; debranching enzymes; endosperm; forage grasses; Poaceae; starch; starch synthase; starch branching enzyme 1. Introduction The grasses can rightly be regarded as a cornerstone of human civilization. -

Molecular Control of Oil Metabolism in the Endosperm of Seeds
International Journal of Molecular Sciences Review Molecular Control of Oil Metabolism in the Endosperm of Seeds Romane Miray †, Sami Kazaz † , Alexandra To and Sébastien Baud * Institut Jean-Pierre Bourgin, INRAE, CNRS, AgroParisTech, Université Paris-Saclay, 78000 Versailles, France; [email protected] (R.M.); [email protected] (S.K.); [email protected] (A.T.) * Correspondence: [email protected] † These authors contributed equally to this work. Abstract: In angiosperm seeds, the endosperm develops to varying degrees and accumulates different types of storage compounds remobilized by the seedling during early post-germinative growth. Whereas the molecular mechanisms controlling the metabolism of starch and seed-storage proteins in the endosperm of cereal grains are relatively well characterized, the regulation of oil metabolism in the endosperm of developing and germinating oilseeds has received particular attention only more recently, thanks to the emergence and continuous improvement of analytical techniques allowing the evaluation, within a spatial context, of gene activity on one side, and lipid metabolism on the other side. These studies represent a fundamental step toward the elucidation of the molecular mechanisms governing oil metabolism in this particular tissue. In particular, they highlight the importance of endosperm-specific transcriptional controls for determining original oil compositions usually observed in this tissue. In the light of this research, the biological functions of oils stored in the endosperm of seeds then appear to be more diverse than simply constituting a source of carbon made available for the germinating seedling. Keywords: seed; endosperm; oil; fatty acid; metabolism Citation: Miray, R.; Kazaz, S.; To, A.; Baud, S. -

Germination : Seeds Come Equipped with Everything Needed to Create A
Germination : Seeds come equipped with everything needed to create a new plant. They’re simply waiting for optimal environmental conditions to be met, then the process of germination can begin. First, the seed absorbs water. The seed coat softens and enzymes within are dissolved. Warm temperatures activate the enzymes to begin working. Oxygen combines with the seed’s stored energy and new tissue is formed through cell elongation and cell division. This description of germination, though over- simplified, gives us a useful glimpse of plant life in its infant stage. · All seed germination involves water, temperature and oxygen. However, each plant species has unique requirements for these three conditions. Therefore, follow seed package instructions carefully. Days to germination and planting depth are important knowledge for successful gardening. · Seeds are often started indoors in order to extend the growing season in our Midwest location. Seed starting in early spring also gives the gardener an opportunity to optimize conditions for germination. Soil temperature can be controlled through the use of heating mats. Watering from the bottom at appropriate intervals keeps the soil moist, not water logged. · Moisture is a pre-requisite for successful germination. However, too much water will exclude oxygen from the seed. Note that seeds need oxygen during this stage of their development; the need for carbon dioxide increases later when leaves emerge and photosynthesis begins. · Each species has an optimal temperature and a range of temperatures for germination. ISU Publication PM 874 “Starting Garden Transplants at Home” includes a table of best soil temperature for germination of several flower and vegetable species.