High-Throughput Proteomic Profiling of the Fish Liver Following Bacterial
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Complement Component 3 (C3) Concentrations in Cancer Patients: a Systematic Review and Meta-Analysis
Complement Component 3 (C3) Concentrations in Cancer Patients: A Systematic Review and Meta-Analysis Zipeng Yang South China Agricultural University Zi-Guo Yuan ( [email protected] ) Anqun Yang Dapeng New District Nan'ao Peolple's Hospital of Shenzhen, Guangdong Province, China Xiu-Xiang Zhang South China Agricultural University College of Agriculture Xiaohu Wang Guangdong Academy of Agricultural Sciences, Guangzhou, Guangdong Province Miao Yu Dongfeng Xiyuan District, Stomatological Hospital Aliated to Guangzhou Medical University, Guangzhou Yasser S.Mahmmod Zagazig University Jorge A Hernandez University of Florida Zhaowen Ren South China Agricultural University Xirui Zhang South China Agricultural University Wei Cong Shandong University Research article Keywords: Complement Component 3, Cancer Patients, Meta-Analysis Posted Date: March 26th, 2020 DOI: https://doi.org/10.21203/rs.3.rs-18256/v1 License: This work is licensed under a Creative Commons Attribution 4.0 International License. Read Full License Page 1/17 Abstract Background The aim of this study was to investigate the association between the serum complement component 3 “C3” level and the patients with different types of cancer. Our study nding would ultimately provide reliable scientic conclusions to guide clinical practice. Methods PubMed, Embase, The Chorane Library and Google Scholar were systematically searched to identify all studies on serum C3 concentrations in cancer patients published as of September 2019. Additionally, we conducted a clinical study on serum C3 in lung cancer patients and healthy people. The levels of serum complement C3 in 84 lung cancer patients and 30 healthy people were examined by ELISA. We used standardized mean differences (SMD) to report the pooled estimation, and I² statistics were calculated to examine the heterogeneity. -

Complement Peptide C3a Receptor 1 Promotes Optic Nerve Degeneration in DBA/2J Mice Jeffrey M
Harder et al. Journal of Neuroinflammation (2020) 17:336 https://doi.org/10.1186/s12974-020-02011-z RESEARCH Open Access Complement peptide C3a receptor 1 promotes optic nerve degeneration in DBA/2J mice Jeffrey M. Harder1, Pete A. Williams1,2 , Catherine E. Braine1,3, Hongtian S. Yang1, Jocelyn M. Thomas1, Nicole E. Foxworth1, Simon W. M. John1,4,5* and Gareth R. Howell1,6,7* Abstract Background: The risk of glaucoma increases significantly with age and exposure to elevated intraocular pressure, two factors linked with neuroinflammation. The complement cascade is a complex immune process with many bioactive end-products, including mediators of inflammation. Complement cascade activation has been shown in glaucoma patients and models of glaucoma. However, the function of complement-mediated inflammation in glaucoma is largely untested. Here, the complement peptide C3a receptor 1 was genetically disrupted in DBA/2J mice, an ocular hypertensive model of glaucoma, to test its contribution to neurodegeneration. Methods: A null allele of C3ar1 was backcrossed into DBA/2J mice. Development of iris disease, ocular hypertension, optic nerve degeneration, retinal ganglion cell activity, loss of RGCs, and myeloid cell infiltration in C3ar1-deficient and sufficient DBA/2J mice were compared across multiple ages. RNA sequencing was performed on microglia from primary culture to determine global effects of C3ar1 on microglia gene expression. Results: Deficiency in C3ar1 lowered the risk of degeneration in ocular hypertensive mice without affecting intraocular pressure elevation at 10.5 months of age. Differences were found in the percentage of mice affected, but not in individual characteristics of disease progression. -

WO 2016/147053 Al 22 September 2016 (22.09.2016) P O P C T
(12) INTERNATIONAL APPLICATION PUBLISHED UNDER THE PATENT COOPERATION TREATY (PCT) (19) World Intellectual Property Organization International Bureau (10) International Publication Number (43) International Publication Date WO 2016/147053 Al 22 September 2016 (22.09.2016) P O P C T (51) International Patent Classification: (71) Applicant: RESVERLOGIX CORP. [CA/CA]; 300, A61K 31/551 (2006.01) A61P 37/02 (2006.01) 4820 Richard Road Sw, Calgary, AB, T3E 6L1 (CA). A61K 31/517 (2006.01) C07D 239/91 (2006.01) (72) Inventors: WASIAK, Sylwia; 431 Whispering Water (21) International Application Number: Trail, Calgary, AB, T3Z 3V1 (CA). KULIKOWSKI, PCT/IB20 16/000443 Ewelina, B.; 31100 Swift Creek Terrace, Calgary, AB, T3Z 0B7 (CA). HALLIDAY, Christopher, R.A.; 403 (22) International Filing Date: 138-18th Avenue SE, Calgary, AB, T2G 5P9 (CA). GIL- 10 March 2016 (10.03.2016) HAM, Dean; 249 Scenic View Close NW, Calgary, AB, (25) Filing Language: English T3L 1Y5 (CA). (26) Publication Language: English (81) Designated States (unless otherwise indicated, for every kind of national protection available): AE, AG, AL, AM, (30) Priority Data: AO, AT, AU, AZ, BA, BB, BG, BH, BN, BR, BW, BY, 62/132,572 13 March 2015 (13.03.2015) US BZ, CA, CH, CL, CN, CO, CR, CU, CZ, DE, DK, DM, 62/264,768 8 December 2015 (08. 12.2015) US DO, DZ, EC, EE, EG, ES, FI, GB, GD, GE, GH, GM, GT, [Continued on nextpage] (54) Title: COMPOSITIONS AND THERAPEUTIC METHODS FOR THE TREATMENT OF COMPLEMENT-ASSOCIATED DISEASES (57) Abstract: The invention comprises methods of modulating the complement cascade in a mammal and for treating and/or preventing diseases and disorders as sociated with the complement pathway by administering a compound of Formula I or Formula II, such as, for example, 2-(4-(2-hydroxyethoxy)-3,5-dimethylphenyl)- 5,7-dimethoxyquinazolin-4(3H)-one or a pharmaceutically acceptable salt thereof. -

Active Complement Component 1, Q
APD208Hu01 200µg Active Complement Component 1, Q Subcomponent B (C1qB) Organism Species: Homo sapiens (Human) Instruction manual FOR RESEARCH USE ONLY NOT FOR USE IN CLINICAL DIAGNOSTIC PROCEDURES 1th Edition (Apr, 2016) [ PROPERTIES ] Source: Prokaryotic expression. Host: E. coli Residues: Gln28~Ala253 Tags: Two N-terminal Tags, His-tag and SUMO-tag Purity: >94% Endotoxin Level: <1.0EU per 1μg (determined by the LAL method). Buffer Formulation: 20mM Tris, 150mM NaCl, pH8.0, containing 0.05% sarcosyl and 5% trehalose. Applications: Cell culture; Activity Assays. (May be suitable for use in other assays to be determined by the end user.) Predicted isoelectric point: 8.9 Predicted Molecular Mass: 37.5kDa Accurate Molecular Mass: 38kDa as determined by SDS-PAGE reducing conditions. [ USAGE ] Reconstitute in 20mM Tris, 150mM NaCl (pH8.0) to a concentration of 0.1-1.0 mg/mL. Do not vortex. [ STORAGE AND STABILITY ] Storage: Avoid repeated freeze/thaw cycles. Store at 2-8oC for one month. Aliquot and store at -80oC for 12 months. Stability Test: The thermal stability is described by the loss rate. The loss rate was determined by accelerated thermal degradation test, that is, incubate the protein at 37oC for 48h, and no obvious degradation and precipitation were observed. The loss rate is less than 5% within the expiration date under appropriate storage condition. [ SEQUENCE ] [ ACTIVITY ] The complement component 1q (or simply C1q) is a protein complex involved in the complement system, which is part of the innate immune system. C1q together with C1r and C1s form the C1 complex. Antibodies of the adaptive immune system can bind antigen, forming an antigen-antibody complex. -

Important Roles of C5a and C5ar in Tumor Development and Cancer Treatment
E3S Web o f Conferences 136, 06012 (2019) https://doi.org/10.1051/e3sconf/20191360 6012 ICBTE 2019 Important roles of C5a and C5aR in tumor development and cancer treatment Wang Yuxuan Bioengineering, School of Life Science, Anhui University,230601 Abstract: The complement system is part of the body's innate defense immune system, which can identify and eliminate invasive pathogenic microorganisms to maintain normal life activities. Complement Component 5a (C5a) is an active anaphylatoxin produced after complement system activation, closely related to tumor formation. C5a is highly expressed in a variety of tumors, and combines with its Complement Component 5a Receptor (C5aR) to increase the proliferation and migration of tumor cells. This review will comprehensively elaborate the important role of C5a/C5aR in the process of tumor genesis and development from the three aspects of signal transduction pathways related to tumor, C5a/C5aR and tumor formation, and C5a/C5aR inhibitors and tumor therapy. Finally, the principle of complement inhibition is used to inhibit tumor metastasis, reduce the rate of tumor diffusion, and control the trend of tumor deterioration. 1 Introduction The generation of tumor is closely related to the microenvironment in which tumor is located. A variety of cells in the microenvironment, such as lymphocytes, macrophages, fibroblasts and various molecules, such as complement and cytokines, are involved in the survival, proliferation and metastasis of tumor cells. Complement Component is a protein with enzyme activity. Since the discovery of complement system by Belgian doctor J. Bordet in 1890, a large number of studies have shown that complement exists in human serum, tissue fluid and cell membrane surface1. -

Shiga Toxin 2A Binds to Complement Components C3b and C5 and Upregulates Their Gene Expression in Human Cell Lines
toxins Article Shiga Toxin 2a Binds to Complement Components C3b and C5 and Upregulates Their Gene Expression in Human Cell Lines Sára Kellnerová 1,†, Sneha Chatterjee 1,†, Rafael Bayarri-Olmos 2 , Louise Justesen 2 , Heribert Talasz 3, Wilfried Posch 1 , Samyr Kenno 1, Peter Garred 2, Dorothea Orth-Höller 1,*, Marco Grasse 1 and Reinhard Würzner 1,* 1 Institute of Hygiene and Medical Microbiology, Medical University of Innsbruck, Schöpfstraβe 41, A-6020 Innsbruck, Austria; [email protected] (S.K.); [email protected] (S.C.); [email protected] (W.P.); [email protected] (S.K.); [email protected] (M.G.) 2 Laboratory of Molecular Medicine, Department of Clinical Immunology Section 7631, Rigshospitalet, University of Copenhagen, Ole Maaloesvej 26, 2200 Copenhagen, Denmark; [email protected] (R.B.-O.); [email protected] (L.J.); [email protected] (P.G.) 3 Centre of Chemistry and Biomedicine, Division of Clinical Biochemistry, Medical University of Innsbruck, Innrain 80, A-6020 Innsbruck, Austria; [email protected] * Correspondence: [email protected] (D.O.-H.); [email protected] (R.W.); Tel.: +43-512-900-370-707 (R.W.) † First authors. Abstract: Enterohemorrhagic Escherichia coli (EHEC) infections can cause EHEC-associated hemolytic uremic syndrome (eHUS) via its main virulent factor, Shiga toxins (Stxs). Complement has been reported to be involved in the progression of eHUS. The aim of this study was to investigate the interactions of the most effective subtype of the toxin, Stx2a, with pivotal complement proteins C3b and C5. -

Complement Component 4 Genes Contribute Sex-Specific Vulnerability in Diverse Illnesses
bioRxiv preprint doi: https://doi.org/10.1101/761718; this version posted September 9, 2019. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-ND 4.0 International license. Complement component 4 genes contribute sex-specific vulnerability in diverse illnesses Nolan Kamitaki1,2, Aswin Sekar1,2, Robert E. Handsaker1,2, Heather de Rivera1,2, Katherine Tooley1,2, David L. Morris3, Kimberly E. Taylor4, Christopher W. Whelan1,2, Philip Tombleson3, Loes M. Olde Loohuis5,6, Schizophrenia Working Group of the Psychiatric Genomics Consortium7, Michael Boehnke8, Robert P. Kimberly9, Kenneth M. Kaufman10, John B. Harley10, Carl D. Langefeld11, Christine E. Seidman1,12,13, Michele T. Pato14, Carlos N. Pato14, Roel A. Ophoff5,6, Robert R. Graham15, Lindsey A. Criswell4, Timothy J. Vyse3, Steven A. McCarroll1,2 1 Department of Genetics, Harvard Medical School, Boston, Massachusetts 02115, USA 2 Stanley Center for Psychiatric Research, Broad Institute of MIT and Harvard, Cambridge, Massachusetts 02142, USA 3 Department of Medical and Molecular Genetics, King’s College London, London WC2R 2LS, UK 4 Rosalind Russell / Ephraim P Engleman Rheumatology Research Center, Division of Rheumatology, UCSF School of Medicine, San Francisco, California 94143, USA 5 Department of Human Genetics, David Geffen School of Medicine, University of California, Los Angeles, California 90095, USA 6 Center for Neurobehavioral Genetics, Semel Institute for Neuroscience and Human Behavior, University of California, Los Angeles, California 90095, USA 7 A full list of collaborators is in Supplementary Information. -

Review Article Complement System and Age-Related Macular Degeneration: Implications of Gene-Environment Interaction for Preventive and Personalized Medicine
Hindawi BioMed Research International Volume 2018, Article ID 7532507, 13 pages https://doi.org/10.1155/2018/7532507 Review Article Complement System and Age-Related Macular Degeneration: Implications of Gene-Environment Interaction for Preventive and Personalized Medicine Andrea Maugeri ,1 Martina Barchitta ,1 Maria Grazia Mazzone,2 Francesco Giuliano,2 and Antonella Agodi 1 1 Department of Medical and Surgical Sciences and Advanced Technologies “GF Ingrassia”, University of Catania, Via S. Sofa 87, 95123 Catania, Italy 2SIFI SpA, Research and Development Department, Via Ercole Patti 36, 95025 Catania, Italy Correspondence should be addressed to Antonella Agodi; [email protected] Received 18 May 2018; Accepted 18 July 2018; Published 26 August 2018 Academic Editor: Sajib Chakraborty Copyright © 2018 Andrea Maugeri et al. Tis is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Age-related macular degeneration (AMD) is the most common cause of visual loss in developed countries, with a signifcant economic and social burden on public health. Although genome-wide and gene-candidate studies have been enabled to identify genetic variants in the complement system associated with AMD pathogenesis, the efect of gene-environment interaction is still under debate. In this review we provide an overview of the role of complement system and its genetic variants in AMD, summarizing the consequences of the interaction between genetic and environmental risk factors on AMD onset, progression, and therapeutic response. Finally, we discuss the perspectives of current evidence in the feld of genomics driven personalized medicine and public health. -

Supplementary Table S4. FGA Co-Expressed Gene List in LUAD
Supplementary Table S4. FGA co-expressed gene list in LUAD tumors Symbol R Locus Description FGG 0.919 4q28 fibrinogen gamma chain FGL1 0.635 8p22 fibrinogen-like 1 SLC7A2 0.536 8p22 solute carrier family 7 (cationic amino acid transporter, y+ system), member 2 DUSP4 0.521 8p12-p11 dual specificity phosphatase 4 HAL 0.51 12q22-q24.1histidine ammonia-lyase PDE4D 0.499 5q12 phosphodiesterase 4D, cAMP-specific FURIN 0.497 15q26.1 furin (paired basic amino acid cleaving enzyme) CPS1 0.49 2q35 carbamoyl-phosphate synthase 1, mitochondrial TESC 0.478 12q24.22 tescalcin INHA 0.465 2q35 inhibin, alpha S100P 0.461 4p16 S100 calcium binding protein P VPS37A 0.447 8p22 vacuolar protein sorting 37 homolog A (S. cerevisiae) SLC16A14 0.447 2q36.3 solute carrier family 16, member 14 PPARGC1A 0.443 4p15.1 peroxisome proliferator-activated receptor gamma, coactivator 1 alpha SIK1 0.435 21q22.3 salt-inducible kinase 1 IRS2 0.434 13q34 insulin receptor substrate 2 RND1 0.433 12q12 Rho family GTPase 1 HGD 0.433 3q13.33 homogentisate 1,2-dioxygenase PTP4A1 0.432 6q12 protein tyrosine phosphatase type IVA, member 1 C8orf4 0.428 8p11.2 chromosome 8 open reading frame 4 DDC 0.427 7p12.2 dopa decarboxylase (aromatic L-amino acid decarboxylase) TACC2 0.427 10q26 transforming, acidic coiled-coil containing protein 2 MUC13 0.422 3q21.2 mucin 13, cell surface associated C5 0.412 9q33-q34 complement component 5 NR4A2 0.412 2q22-q23 nuclear receptor subfamily 4, group A, member 2 EYS 0.411 6q12 eyes shut homolog (Drosophila) GPX2 0.406 14q24.1 glutathione peroxidase -

Deficiency of Complement Component 5 Ameliorates Glaucoma in DBA/2J Mice
Rowan University Rowan Digital Works Faculty Scholarship for the College of Science & Mathematics College of Science & Mathematics 6-27-2013 Deficiency of complement component 5 ameliorates glaucoma in DBA/2J mice Gareth R. Howell Ileana Soto Reyes Rowan University, [email protected] Margaret Ryan Leah C. Graham Richard S. Smith See next page for additional authors Follow this and additional works at: https://rdw.rowan.edu/csm_facpub Part of the Neuroscience and Neurobiology Commons Recommended Citation Howell, G. R., Soto, I., Ryan, M., Graham, L. C., Smith, R. S., & John, S. W. (2013). Deficiency of complement component 5 ameliorates glaucoma in DBA/2J mice. Journal of Neuroinflammation, 10, 76-2094-10-76. This Article is brought to you for free and open access by the College of Science & Mathematics at Rowan Digital Works. It has been accepted for inclusion in Faculty Scholarship for the College of Science & Mathematics by an authorized administrator of Rowan Digital Works. Authors Gareth R. Howell, Ileana Soto Reyes, Margaret Ryan, Leah C. Graham, Richard S. Smith, and Simon W.M. John This article is available at Rowan Digital Works: https://rdw.rowan.edu/csm_facpub/24 Howell et al. Journal of Neuroinflammation 2013, 10:76 JOURNAL OF http://www.jneuroinflammation.com/content/10/1/76 NEUROINFLAMMATION RESEARCH Open Access Deficiency of complement component 5 ameliorates glaucoma in DBA/2J mice Gareth R Howell1*†, Ileana Soto1†, Margaret Ryan1, Leah C Graham1, Richard S Smith1 and Simon WM John1,2,3* Abstract Background: Glaucoma is an age-related neurodegenerative disorder involving the loss of retinal ganglion cells (RGCs), which results in blindness. -
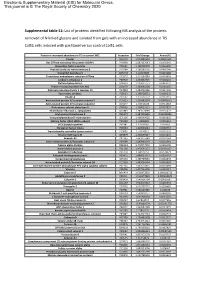
Supplemental Table S1
Electronic Supplementary Material (ESI) for Molecular Omics. This journal is © The Royal Society of Chemistry 2020 Supplemental table S1: List of proteins identified following MS analysis of the proteins removed of N-linked glycans and isolated from gels with an increased abundance in TIS Cal51 cells induced with paclitaxel versus control Cal51 cells. Protein in increased abundance in TIS vs control WCL Accession Fold Change Anova (P) Plectin Q15149 1.073855593 0.00691631 Ras GTPase-activating-like protein IQGAP1 P46940 1.087337643 0.0176342 Elongation factor1-gamma P26641 1.138709703 0.0116496 Peptidyl-prolyl cis-transisomerase B P23284 1.188383105 0.0436246 Dipeptidyl peptidase 3 Q9NY33 1.20163605 0.0215448 Transitional endoplasmic reticulum ATPase P55072 1.214194884 0.0449691 Carbonic anhydrase 2 P00918 1.232852325 0.0158141 Clathrin heavy chain 1 Q00610 1.239621773 0.0463237 Protein transport protein Sec 31A O94979 1.263565104 0.0284155 Aldo-ketoreductase family 1 member C1 Q04828 1.282092186 0.0324406 Spermidine synthase P19623 1.298728621 0.0196232 Plastin-3 P13797 1.310756772 0.0161319 Actin-related protein 2/3 complex subunit 5 O15511 1.333483524 0.00476923 Actin-related protein 2/3 complex subunit 2 O15144 1.35416168 0.0411018 Proteasome subunit alpha type-5 P28066 1.358015551 0.0337657 Thioredoxin reductase 1, cytoplasmic Q16881 1.383670089 0.0235472 Acyl-protein thioesterase 2 O95372 1.387415589 0.00233899 Isoaspartylpeptidase/L-asparaginase Q7L266 1.408149002 0.0319602 Splicing factor U2AF 65kDa subunit P26368 1.41489991 0.0256619 -

In Kidney Injury
antioxidants Review Regulation of Complement Activation by Heme Oxygenase-1 (HO-1) in Kidney Injury Maria G. Detsika 1,* and Elias A. Lianos 2,3 1 First Department of Critical Care Medicine & Pulmonary Services, GP Livanos and M. Simou Laboratories, National & Kapodistrian University of Athens, Medical School, Evangelismos Hospital, 10675 Athens, Greece 2 Thorax Foundation, Research Center of Intensive Care and Emergency Thoracic Medicine, 10675 Athens, Greece; [email protected] 3 Veterans Affairs Medical Center and Virginia Tech, Carilion School of Medicine, 1970 Roanoke Blvd, Salem, VA 24153, USA * Correspondence: [email protected]; Tel.: +30-210-723552; Fax: +30-210-7239127 Abstract: Heme oxygenase is a cytoprotective enzyme with strong antioxidant and anti-apoptotic properties. Its cytoprotective role is mainly attributed to its enzymatic activity, which involves the degradation of heme to biliverdin with simultaneous release of carbon monoxide (CO). Recent studies uncovered a new cytoprotective role for heme oxygenase-1 (HO-1) by identifying a regulatory role on the complement control protein decay-accelerating factor. This is a key complement regulatory protein preventing dysregulation or overactivation of complement cascades that can cause kidney injury. Cell-specific targeting of HO-1 induction may, therefore, be a novel approach to attenuate complement-dependent forms of kidney disease. Keywords: heme; heme oxygenase-1 (HO-1); complement; kidney injury 1. Introduction Citation: Detsika, M.G.; Lianos, E.A. Although the role of heme, in various cellular processes, such as gene transcription Regulation of Complement and translation and cellular differentiation, proliferation, and apoptosis, has been known Activation by Heme Oxygenase-1 for decades, the role of the heme-degrading enzyme heme oxygenase-1 (HO-1) only gained (HO-1) in Kidney Injury.