Chlamydia Trachomatis Infection: Host Immune Responses and Potential Vaccines
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Compendium of Measures to Control Chlamydia Psittaci Infection Among
Compendium of Measures to Control Chlamydia psittaci Infection Among Humans (Psittacosis) and Pet Birds (Avian Chlamydiosis), 2017 Author(s): Gary Balsamo, DVM, MPH&TMCo-chair Angela M. Maxted, DVM, MS, PhD, Dipl ACVPM Joanne W. Midla, VMD, MPH, Dipl ACVPM Julia M. Murphy, DVM, MS, Dipl ACVPMCo-chair Ron Wohrle, DVM Thomas M. Edling, DVM, MSpVM, MPH (Pet Industry Joint Advisory Council) Pilar H. Fish, DVM (American Association of Zoo Veterinarians) Keven Flammer, DVM, Dipl ABVP (Avian) (Association of Avian Veterinarians) Denise Hyde, PharmD, RP Preeta K. Kutty, MD, MPH Miwako Kobayashi, MD, MPH Bettina Helm, DVM, MPH Brit Oiulfstad, DVM, MPH (Council of State and Territorial Epidemiologists) Branson W. Ritchie, DVM, MS, PhD, Dipl ABVP, Dipl ECZM (Avian) Mary Grace Stobierski, DVM, MPH, Dipl ACVPM (American Veterinary Medical Association Council on Public Health and Regulatory Veterinary Medicine) Karen Ehnert, and DVM, MPVM, Dipl ACVPM (American Veterinary Medical Association Council on Public Health and Regulatory Veterinary Medicine) Thomas N. Tully JrDVM, MS, Dipl ABVP (Avian), Dipl ECZM (Avian) (Association of Avian Veterinarians) Source: Journal of Avian Medicine and Surgery, 31(3):262-282. Published By: Association of Avian Veterinarians https://doi.org/10.1647/217-265 URL: http://www.bioone.org/doi/full/10.1647/217-265 BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use. -

Chlamydia Cell Biology and Pathogenesis
HHS Public Access Author manuscript Author ManuscriptAuthor Manuscript Author Nat Rev Manuscript Author Microbiol. Author Manuscript Author manuscript; available in PMC 2016 June 01. Published in final edited form as: Nat Rev Microbiol. 2016 June ; 14(6): 385–400. doi:10.1038/nrmicro.2016.30. Chlamydia cell biology and pathogenesis Cherilyn Elwell, Kathleen Mirrashidi, and Joanne Engel Departments of Medicine, Microbiology and Immunology, University of California, San Francisco, California 94143, USA Abstract Chlamydia spp. are important causes of human disease for which no effective vaccine exists. These obligate intracellular pathogens replicate in a specialized membrane compartment and use a large arsenal of secreted effectors to survive in the hostile intracellular environment of the host. In this Review, we summarize the progress in decoding the interactions between Chlamydia spp. and their hosts that has been made possible by recent technological advances in chlamydial proteomics and genetics. The field is now poised to decipher the molecular mechanisms that underlie the intimate interactions between Chlamydia spp. and their hosts, which will open up many exciting avenues of research for these medically important pathogens. Chlamydiae are Gram-negative, obligate intracellular pathogens and symbionts of diverse 1 organisms, ranging from humans to amoebae . The best-studied group in the Chlamydiae phylum is the Chlamydiaceae family, which comprises 11 species that are pathogenic to 1 humans or animals . Some species that are pathogenic to animals, such as the avian 1 2 pathogen Chlamydia psittaci, can be transmitted to humans , . The mouse pathogen 3 Chlamydia muridarum is a useful model of genital tract infections . Chlamydia trachomatis and Chlamydia pneumoniae, the major species that infect humans, are responsible for a wide 2 4 range of diseases , and will be the focus of this Review. -

Chlamydia Trachomatis Infection Is Driven by Nonprotective Immune Cells That Are Distinct from Protective Populations
Pathology after Chlamydia trachomatis infection is driven by nonprotective immune cells that are distinct from protective populations Rebeccah S. Lijeka,b,1, Jennifer D. Helblea, Andrew J. Olivea,c, Kyra W. Seigerb, and Michael N. Starnbacha,1 aDepartment of Microbiology and Immunobiology, Harvard Medical School, Boston, MA 02115; bDepartment of Biological Sciences, Mount Holyoke College, South Hadley, MA 01075; and cDepartment of Microbiology and Physiological Systems, University of Massachusetts Medical School, Worcester, MA 01605 Edited by Rafi Ahmed, Emory University, Atlanta, GA, and approved December 27, 2017 (received for review June 23, 2017) Infection with Chlamydia trachomatis drives severe mucosal immu- sequence identity, Chlamydia muridarum, the extent to which the nopathology; however, the immune responses that are required for molecular pathogenesis of C. muridarum represents that of Ct is mediating pathology vs. protection are not well understood. Here, unknown (6). Ct serovar L2 (Ct L2) is capable of infecting the we employed a mouse model to identify immune responses re- mouse upper genital tract when inoculated across the cervix into quired for C. trachomatis-induced upper genital tract pathology the uterus (7, 8) but it does not induce robust immunopathology. and to determine whether these responses are also required for This is consistent with the human disease phenotype caused by Ct L2, bacterial clearance. In mice as in humans, immunopathology was which disseminates to the lymph nodes causing lymphogranuloma characterized by extravasation of leukocytes into the upper genital venereum (LGV) and is not a major cause of mucosal immunopa- thology in the female upper genital tract (uterus and ovaries). tract that occluded luminal spaces in the uterus and ovaries. -

Table S4. Phylogenetic Distribution of Bacterial and Archaea Genomes in Groups A, B, C, D, and X
Table S4. Phylogenetic distribution of bacterial and archaea genomes in groups A, B, C, D, and X. Group A a: Total number of genomes in the taxon b: Number of group A genomes in the taxon c: Percentage of group A genomes in the taxon a b c cellular organisms 5007 2974 59.4 |__ Bacteria 4769 2935 61.5 | |__ Proteobacteria 1854 1570 84.7 | | |__ Gammaproteobacteria 711 631 88.7 | | | |__ Enterobacterales 112 97 86.6 | | | | |__ Enterobacteriaceae 41 32 78.0 | | | | | |__ unclassified Enterobacteriaceae 13 7 53.8 | | | | |__ Erwiniaceae 30 28 93.3 | | | | | |__ Erwinia 10 10 100.0 | | | | | |__ Buchnera 8 8 100.0 | | | | | | |__ Buchnera aphidicola 8 8 100.0 | | | | | |__ Pantoea 8 8 100.0 | | | | |__ Yersiniaceae 14 14 100.0 | | | | | |__ Serratia 8 8 100.0 | | | | |__ Morganellaceae 13 10 76.9 | | | | |__ Pectobacteriaceae 8 8 100.0 | | | |__ Alteromonadales 94 94 100.0 | | | | |__ Alteromonadaceae 34 34 100.0 | | | | | |__ Marinobacter 12 12 100.0 | | | | |__ Shewanellaceae 17 17 100.0 | | | | | |__ Shewanella 17 17 100.0 | | | | |__ Pseudoalteromonadaceae 16 16 100.0 | | | | | |__ Pseudoalteromonas 15 15 100.0 | | | | |__ Idiomarinaceae 9 9 100.0 | | | | | |__ Idiomarina 9 9 100.0 | | | | |__ Colwelliaceae 6 6 100.0 | | | |__ Pseudomonadales 81 81 100.0 | | | | |__ Moraxellaceae 41 41 100.0 | | | | | |__ Acinetobacter 25 25 100.0 | | | | | |__ Psychrobacter 8 8 100.0 | | | | | |__ Moraxella 6 6 100.0 | | | | |__ Pseudomonadaceae 40 40 100.0 | | | | | |__ Pseudomonas 38 38 100.0 | | | |__ Oceanospirillales 73 72 98.6 | | | | |__ Oceanospirillaceae -

Product Description EN Bactoreal® Kit Chlamydiaceae
Product Description BactoReal® Kit Chlamydiaceae For research only, not for diagnostic use BactoReal® Kit Chlamydiaceae Order no. Reactions Pathogen Internal positive control DVEB03113 100 FAM channel Cy5 channel DVEB03153 50 FAM channel Cy5 channel DVEB03111 100 FAM channel VIC/HEX channel DVEB03151 50 FAM channel VIC/HEX channel Kit contents: Detection assay for Chlamydia and Chlamydophila species Detection assay for internal positive control (control of amplification) DNA reaction mix (contains uracil-N glycosylase, UNG) Positive control for Chlamydiaceae Water Background: The Chlamydiaceae family currently includes two genera and one candidate genus: genus Chlamydia (including the species Chlamydia muridarum, Chlamydia suis, Chlamydia trachomatis), genus Chlamydophila (including the species Chlamydophila abortus, Chlamydophila caviae Chlamydophila felis, Chlamydia pecorum, Chlamydophila pneumoniae, Chlamydophila psittaci), and candidatus Clavochlamydia. All Chlamydiaceae are obligate intracellular Gram-negative bacteria that cause diseases in humans and animals worldwide. Description: BactoReal® Kit Chlamydiaceae is based on the amplification and detection of the 23S rRNA gene of C. muridarum, C. suis, C. trachomatis, C. abortus, C. caviae, C. felis, C. pecorum, C. pneumoniae and C. psittaci using real-time PCR. It allows the rapid and sensitive detection of the 23S rRNA gene of Chlamydiaceae from DNA samples purified from different sample material (e.g. with the QIAamp DNA Mini Kit). For subtyping please contact ingenetix. PCR-platforms: BactoReal® Kit Chlamydiaceae is developed and validated for the ABI PRISM® 7500 instrument (Life Technologies), LightCycler® 480 (Roche) and Mx3005P® QPCR System (Agilent), but is also suitable for other real-time PCR instruments. Sensitivity and specificity: BactoReal® Kit Chlamydiaceae detects at least 10 copies/reaction. -

Leptospirosis: a Waterborne Zoonotic Disease of Global Importance
August 2006 volume 22 number 08 Leptospirosis: A waterborne zoonotic disease of global importance INTRODUCTION syndrome has two phases: a septicemic and an immune phase (Levett, 2005). Leptospirosis is considered one of the most common zoonotic diseases It is in the immune phase that organ-specific damage and more severe illness globally. In the United States, outbreaks are increasingly being reported is seen. See text box for more information on the two phases. The typical among those participating in recreational water activities (Centers for Disease presenting signs of leptospirosis in humans are fever, headache, chills, con- Control and Prevention [CDC], 1996, 1998, and 2001) and sporadic cases are junctival suffusion, and myalgia (particularly in calf and lumbar areas) often underdiagnosed. With the onset of warm temperatures, increased (Heymann, 2004). Less common signs include a biphasic fever, meningitis, outdoor activities, and travel, Georgia may expect to see more leptospirosis photosensitivity, rash, and hepatic or renal failure. cases. DIAGNOSIS OF LEPTOSPIROSIS Leptospirosis is a zoonosis caused by infection with the bacterium Leptospira Detecting serum antibodies against leptospira interrogans. The disease occurs worldwide, but it is most common in temper- • Microscopic Agglutination Titers (MAT) ate regions in the late summer and early fall and in tropical regions during o Paired serum samples which show a four-fold rise in rainy seasons. It is not surprising that Hawaii has the highest incidence of titer confirm the diagnosis; a single high titer in a per- leptospirosis in the United States (Levett, 2005). The reservoir of pathogenic son clinically suspected to have leptospirosis is highly leptospires is the renal tubules of wild and domestic animals. -

Gonorrhea, Chlamydia, and Syphilis
2019 GONORRHEA, CHLAMYDIA, AND SYPHILIS AND CHLAMYDIA, GONORRHEA, Dedication TAG would like to thank the National Coalition of STD Directors for funding and input on the report. THE PIPELINE REPORT Pipeline for Gonorrhea, Chlamydia, and Syphilis By Jeremiah Johnson Introduction The current toolbox for addressing gonorrhea, chlamydia, and syphilis is inadequate. At a time where all three epidemics are dramatically expanding in locations all around the globe, including record-breaking rates of new infections in the United States, stakeholders must make do with old tools, inadequate systems for addressing sexual health, and a sparse research pipeline of new treatment, prevention, and diagnostic options. Lack of investment in sexual health research has left the field with inadequate prevention options, and limited access to infrastructure for testing and treatment have allowed sexually transmitted infections (STIs) to flourish. The consequences of this underinvestment are large: according to the World Health Organization (WHO), in 2012 there were an estimated 357 million new infections (roughly 1 million per day) of the four curable STIs: gonorrhea, chlamydia, syphilis, and trichomoniasis.1 In the United States, the three reportable STIs that are the focus of this report—gonorrhea, chlamydia, and syphilis—are growing at record paces. In 2017, a total of 30,644 cases of primary and secondary (P&S) syphilis—the most infectious stages of the disease—were reported in the United States. Since reaching a historic low in 2000 and 2001, the rate of P&S syphilis has increased almost every year, increasing 10.5% during 2016–2017. Also in 2017, 555,608 cases of gonorrhea were reported to the U.S. -
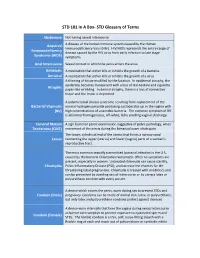
STD Glossary of Terms
STD 101 In A Box- STD Glossary of Terms Abstinence Not having sexual intercourse Acquired A disease of the human immune system caused by the Human Immunodeficiency Virus (HIV). HIV/AIDS represents the entire range of Immunodeficiency disease caused by the HIV virus from early infection to late stage Syndrome (AIDS) symptoms. Anal Intercourse Sexual contact in which the penis enters the anus. Antibiotic A medication that either kills or inhibits the growth of a bacteria. Antiviral A medication that either kills or inhibits the growth of a virus. A thinning of tissue modified by the location. In epidermal atrophy, the epidermis becomes transparent with a loss of skin texture and cigarette Atrophic paper-like wrinkling. In dermal atrophy, there is a loss of connective tissue and the lesion is depressed. A polymicrobial clinical syndrome resulting from replacement of the Bacterial Vaginosis normal hydrogen peroxide producing Lactobacillus sp. in the vagina with (BV) high concentrations of anaerobic bacteria. The common symptom of BV is abnormal homogeneous, off-white, fishy smelling vaginal discharge. Cervical Motion A sign found on pelvic examination suggestive of pelvic pathology; when Tenderness (CMT) movement of the cervix during the bimanual exam elicits pain. The lower, cylindrical end of the uterus that forms a narrow canal Cervix connecting the upper (uterus) and lower (vagina) parts of a woman's reproductive tract. The most common sexually transmitted bacterial infection in the U.S., caused by the bacteria Chlamydia trachomatis. Often no symptoms are present, especially in women. Untreated chlamydia can cause sterility, Chlamydia Pelvic Inflammatory Disease (PID), and increase the chances for life- threatening tubal pregnancies. -

Infection Genital Tract Chlamydia Muridarum Essential for Normal
The Journal of Immunology CD4+ T Cell Expression of MyD88 Is Essential for Normal Resolution of Chlamydia muridarum Genital Tract Infection Lauren C. Frazer,*,† Jeanne E. Sullivan,† Matthew A. Zurenski,† Margaret Mintus,† Tammy E. Tomasak,† Daniel Prantner,‡ Uma M. Nagarajan,† and Toni Darville*,† Resolution of Chlamydia genital tract infection is delayed in the absence of MyD88. In these studies, we first used bone marrow chimeras to demonstrate a requirement for MyD88 expression by hematopoietic cells in the presence of a wild-type epithelium. Using mixed bone marrow chimeras we then determined that MyD88 expression was specifically required in the adaptive immune compartment. Furthermore, adoptive transfer experiments revealed that CD4+ T cell expression of MyD88 was necessary for normal resolution of genital tract infection. This requirement was associated with a reduced ability of MyD882/2CD4+ T cells to accumulate in the draining lymph nodes and genital tract when exposed to the same inflammatory milieu as wild-type CD4+ T cells. We also demonstrated that the impaired infection control we observed in the absence of MyD88 could not be recapitulated by deficiencies in TLR or IL-1R signaling. In vitro, we detected an increased frequency of apoptotic MyD882/2CD4+ T cells upon activation in the absence of exogenous ligands for receptors upstream of MyD88. These data reveal an intrinsic requirement for MyD88 in CD4+ T cells during Chlamydia infection and indicate that the importance of MyD88 extends beyond innate immune responses by directly influencing adaptive immunity. The Journal of Immunology, 2013, 191: 4269–4279. hlamydia trachomatis infections of the female repro- of an adaptive immune response (19), but overly robust innate ductive tract can result in serious pathophysiology in- immune activation results in tissue damage. -

Chlamydia Trachomatis and Chlamydia Pneumoniae Interaction with the Host: Latest Advances and Future Prospective
microorganisms Review Chlamydia trachomatis and Chlamydia pneumoniae Interaction with the Host: Latest Advances and Future Prospective Marisa Di Pietro 1,* , Simone Filardo 1 , Silvio Romano 2 and Rosa Sessa 1 1 Department of Public Health and Infectious Diseases, Section of Microbiology, University of Rome “Sapienza”, 00185 Rome, Italy; simone.fi[email protected] (S.F.); [email protected] (R.S.) 2 Cardiology, Department of Life, Health and Environmental Sciences, University of L’Aquila, 67100 L’Aquila, Italy; [email protected] * Correspondence: [email protected] Received: 15 April 2019; Accepted: 14 May 2019; Published: 16 May 2019 Abstract: Research in Chlamydia trachomatis and Chlamydia pneumoniae has gained new traction due to recent advances in molecular biology, namely the widespread use of the metagenomic analysis and the development of a stable genomic transformation system, resulting in a better understanding of Chlamydia pathogenesis. C. trachomatis, the leading cause of bacterial sexually transmitted diseases, is responsible of cervicitis and urethritis, and C. pneumoniae, a widespread respiratory pathogen, has long been associated with several chronic inflammatory diseases with great impact on public health. The present review summarizes the current evidence regarding the complex interplay between C. trachomatis and host defense factors in the genital micro-environment as well as the key findings in chronic inflammatory diseases associated to C. pneumoniae. Keywords: Chlamydia trachomatis; Chlamydia pneumoniae; host-pathogen interaction 1. Introduction Currently, there is a renewed research interest in Chlamydiae that cause a broad spectrum of pathologies of varying severity in human, mainly Chlamydia trachomatis and Chlamydia pneumoniae [1,2]. Advances in molecular biology and, in particular, the recent advent of metagenomic analysis as well as the development of a stable genomic transformation system in Chlamydiae have significantly contributed to expanding our understanding of Chlamydia pathogenesis [3–5]. -

Chlamydial Genital Infection(Chlamydia Trachomatis)
Chlamydial Genital Infection (Chlamydia trachomatis) February 2003 1) THE DISEASE AND ITS EPIDEMIOLOGY A. Etiologic Agent Chlamydial genital infection (CGI) is caused by the obligate, intracellular bacterium Chlamydia trachomatis immunotypes D through K. B. Clinical Description and Laboratory Diagnosis A sexually transmitted genital infection that manifests in males primarily as urethritis and in females as mucopurulent cervicitis. Clinical manifestations are difficult to distinguish from gonorrhea. Males may present with a mucopurulent discharges of scanty to moderate quantity, urethral itching and dysuria. Asymptomatic infection may be found in 1%-25% of sexually active men. Possible complications include epididymitis, infertility and Reiter syndrome. Anorectal intercourse may result in chlamydial proctitis. Women frequently present with a mucopurulent endocervical discharge including edema, erythema and easily induced endocervical bleeding. However, most women with endocervical or urethral infections are asymptomatic. Possible complications include salpingitis with subsequent risk of infertility and ectopic pregnancy. Asymptomatic chronic infections of the endometrium and fallopian tubes may lead to the same outcomes. Less frequent manifestations include bartholinitis, urethral syndrome with dysuria and pyuria, perihepatitis (Fitz-Hugh-Curtis syndrome), and proctitis. Infection during pregnancy may result in premature rupture of membranes and preterm delivery and conjunctival and pneumonic infection of the newborn. Laboratory diagnosis is based upon the identification of Chlamydia in intraurethral or endocervical smear by direct immunofluorescence test, enzyme immunoassay, DNA probe, and nucleic acid amplification test (NAAT) or cell culture. NAAT can be used with urine specimens. C. Vectors and Reservoirs Humans. D. Modes of Transmission By sexual contact and through perinatal exposure to the mother’s infected cervix. -

Chlamydia, Gonorrhoea, Trichomoniasis and Syphilis
Research Chlamydia, gonorrhoea, trichomoniasis and syphilis: global prevalence and incidence estimates, 2016 Jane Rowley,a Stephen Vander Hoorn,b Eline Korenromp,c Nicola Low,d Magnus Unemo,e Laith J Abu- Raddad,f R Matthew Chico,g Alex Smolak,f Lori Newman,h Sami Gottlieb,a Soe Soe Thwin,a Nathalie Brouteta & Melanie M Taylora Objective To generate estimates of the global prevalence and incidence of urogenital infection with chlamydia, gonorrhoea, trichomoniasis and syphilis in women and men, aged 15–49 years, in 2016. Methods For chlamydia, gonorrhoea and trichomoniasis, we systematically searched for studies conducted between 2009 and 2016 reporting prevalence. We also consulted regional experts. To generate estimates, we used Bayesian meta-analysis. For syphilis, we aggregated the national estimates generated by using Spectrum-STI. Findings For chlamydia, gonorrhoea and/or trichomoniasis, 130 studies were eligible. For syphilis, the Spectrum-STI database contained 978 data points for the same period. The 2016 global prevalence estimates in women were: chlamydia 3.8% (95% uncertainty interval, UI: 3.3–4.5); gonorrhoea 0.9% (95% UI: 0.7–1.1); trichomoniasis 5.3% (95% UI:4.0–7.2); and syphilis 0.5% (95% UI: 0.4–0.6). In men prevalence estimates were: chlamydia 2.7% (95% UI: 1.9–3.7); gonorrhoea 0.7% (95% UI: 0.5–1.1); trichomoniasis 0.6% (95% UI: 0.4–0.9); and syphilis 0.5% (95% UI: 0.4–0.6). Total estimated incident cases were 376.4 million: 127.2 million (95% UI: 95.1–165.9 million) chlamydia cases; 86.9 million (95% UI: 58.6–123.4 million) gonorrhoea cases; 156.0 million (95% UI: 103.4–231.2 million) trichomoniasis cases; and 6.3 million (95% UI: 5.5–7.1 million) syphilis cases.