Molecular Network Pathway of ERBB in Diffuse-Type Gastric Cancer
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Mutation Analysis in Myeloproliferative Neoplasms AHS - M2101
Corporate Medical Policy Mutation Analysis in Myeloproliferative Neoplasms AHS - M2101 File Name: mutation_analysis_in_myeloproliferative_neoplasms Origination: 1/1/2019 Last CAP review: 8/2021 Next CAP review: 8/2022 Last Review: 8/2021 Description of Procedure or Service Myeloproliferative neoplasms (MPN) are a heterogeneous group of clonal disorders characterized by overproduction of one or more differentiated myeloid lineages (Grinfeld, Nangalia, & Green, 2017). These include polycythemia vera (PV), essential thrombocythemia (ET), and primary myelofibrosis (PMF). The majority of MPN result from somatic mutations in the 3 driver genes, JAK2, CALR, and MPL, which represent major diagnostic criteria in combination with hematologic and morphological abnormalities (Rumi & Cazzola, 2017). Related Policies: BCR-ABL 1 Testing for Chronic Myeloid Leukemia AHS-M2027 ***Note: This Medical Policy is complex and technical. For questions concerning the technical language and/or specific clinical indications for its use, please consult your physician. Policy BCBSNC will provide coverage for mutation analysis in myeloproliferative neoplasms when it is determined to be medically necessary because the medical criteria and guidelines shown below are met. Benefits Application This medical policy relates only to the services or supplies described herein. Please refer to the Member's Benefit Booklet for availability of benefits. Member's benefits may vary according to benefit design; therefore member benefit language should be reviewed before applying the terms of this medical policy. When Mutation Analysis in Myeloproliferative Neoplasms is covered 1. JAK2, CALR or MPL mutation testing is considered medically necessary for the diagnosis of patients presenting with clinical, laboratory, or pathological findings suggesting classic forms of myeloproliferative neoplasms (MPN), that is, polycythemia vera (PV), essential thrombocythemia (ET), or primary myelofibrosis (PMF) when ordered by a hematology and/or oncology specialist in the following situations: A. -

Human RET Kinase Protein (His Tag)
Human RET Kinase Protein (His Tag) Catalog Number: 11997-H08H1 General Information SDS-PAGE: Gene Name Synonym: CDHF12; CDHR16; HSCR1; MEN2A; MEN2B; MTC1; PTC; RET-ELE1; RET51 Protein Construction: A DNA sequence encoding the extracellular domain of human RET (P07949-1) (Met 1-Arg 635) was fused with a polyhistidine tag at the N- terminus. Source: Human Expression Host: HEK293 Cells QC Testing Purity: > 92 % as determined by SDS-PAGE Protein Description Endotoxin: RET proto-oncogene, also known as RET, is a cell-surface molecule that < 1.0 EU per μg of the protein as determined by the LAL method transduce signals for cell growth and differentiation. It contains 1 cadherin domain and 1 protein kinase domain. RET proto-oncogene belongs to the Stability: protein kinase superfamily, tyr protein kinase family. RET proto-oncogene is involved in numerous cellular mechanisms including cell proliferation, ℃ Samples are stable for up to twelve months from date of receipt at -70 neuronal navigation, cell migration, and cell differentiation upon binding with glial cell derived neurotrophic factor family ligands. It phosphorylates Leu 29 Predicted N terminal: PTK2/FAK1 and regulates both cell death/survival balance and positional Molecular Mass: information. RET is required for the molecular mechanisms orchestration during intestine organogenesis; involved in the development of enteric The recombinant human RET consists of 618 amino acids and has a nervous system and renal organogenesis during embryonic life; promotes calculated molecular mass of 69.1 kDa. The apparent molecular mass of the formation of Peyer's patch-like structures; modulates cell adhesion via the protein is approximately 110-120 kDa in SDS-PAGE under reducing its cleavage; involved in the development of the neural crest. -

Erb‑B2 Receptor Tyrosine Kinase 2 Is Negatively Regulated by the P53‑Responsive Microrna‑3184‑5P in Cervical Cancer Cells
ONCOLOGY REPORTS 45: 95-106, 2021 Erb‑B2 Receptor Tyrosine Kinase 2 is negatively regulated by the p53‑responsive microRNA‑3184‑5p in cervical cancer cells HONGLI LIU1, YUZHI LI1, JING ZHANG1, NAN WU2, FEI LIU2, LIHUA WANG1, YUAN ZHANG1, JING LIU1, XUAN ZHANG3, SUYANG GUO1 and HONGTAO WANG4 Departments of 1Gynecological Oncology and 2Respiration and Anhui Clinical and Preclinical Key Laboratory of Respiratory Disease, First Affiliated Hospital of Bengbu Medical College; Departments of3 Gynecological Oncology and 4Immunology and Anhui Key Laboratory of Infection and Immunity, Bengbu Medical College, Bengbu, Anhui 233030, P.R. China Received November 30, 2019; Accepted October 2, 2020 DOI: 10.3892/or.2020.7862 Abstract. The oncogenic role of Erb-B2 Receptor Tyrosine Introduction Kinase 2 (ERBB2) has been identified in several types of cancer, but less is known on its function and mechanism of Among women, cervical cancer is ranked 4th in global action in cervical cancer cells. The present study employed cancer-associated deaths (1), with over half a million deaths a multipronged approach to investigate the role of ERBB2 in in 2012 (2). Cervical cancer can be broadly categorized into cervical cancer. ERBB2 and microRNA (miR)-3184-5p expres- squamous cell carcinoma, which constitutes the majority of sion was assessed in patient-derived cervical cancer biopsy cases (70-80%) or adenocarcinoma, which comprises 10-15% tissues, revealing that higher levels of ERBB2 and lower levels of cases (3). Cervical cancer is frequently caused by the of miR-3184-5p were associated with clinicopathological indi- oncovirus human papillomavirus (HPV), mainly by types cators of cervical cancer progression. -

Human and Mouse CD Marker Handbook Human and Mouse CD Marker Key Markers - Human Key Markers - Mouse
Welcome to More Choice CD Marker Handbook For more information, please visit: Human bdbiosciences.com/eu/go/humancdmarkers Mouse bdbiosciences.com/eu/go/mousecdmarkers Human and Mouse CD Marker Handbook Human and Mouse CD Marker Key Markers - Human Key Markers - Mouse CD3 CD3 CD (cluster of differentiation) molecules are cell surface markers T Cell CD4 CD4 useful for the identification and characterization of leukocytes. The CD CD8 CD8 nomenclature was developed and is maintained through the HLDA (Human Leukocyte Differentiation Antigens) workshop started in 1982. CD45R/B220 CD19 CD19 The goal is to provide standardization of monoclonal antibodies to B Cell CD20 CD22 (B cell activation marker) human antigens across laboratories. To characterize or “workshop” the antibodies, multiple laboratories carry out blind analyses of antibodies. These results independently validate antibody specificity. CD11c CD11c Dendritic Cell CD123 CD123 While the CD nomenclature has been developed for use with human antigens, it is applied to corresponding mouse antigens as well as antigens from other species. However, the mouse and other species NK Cell CD56 CD335 (NKp46) antibodies are not tested by HLDA. Human CD markers were reviewed by the HLDA. New CD markers Stem Cell/ CD34 CD34 were established at the HLDA9 meeting held in Barcelona in 2010. For Precursor hematopoetic stem cell only hematopoetic stem cell only additional information and CD markers please visit www.hcdm.org. Macrophage/ CD14 CD11b/ Mac-1 Monocyte CD33 Ly-71 (F4/80) CD66b Granulocyte CD66b Gr-1/Ly6G Ly6C CD41 CD41 CD61 (Integrin b3) CD61 Platelet CD9 CD62 CD62P (activated platelets) CD235a CD235a Erythrocyte Ter-119 CD146 MECA-32 CD106 CD146 Endothelial Cell CD31 CD62E (activated endothelial cells) Epithelial Cell CD236 CD326 (EPCAM1) For Research Use Only. -

Wt1) – an Effector in Leukemogenesis?
WILMS’ TUMOUR GENE 1 PROTEIN (WT1) – AN EFFECTOR IN LEUKEMOGENESIS? Vidovic, Karina 2010 Link to publication Citation for published version (APA): Vidovic, K. (2010). WILMS’ TUMOUR GENE 1 PROTEIN (WT1) – AN EFFECTOR IN LEUKEMOGENESIS?. Lund University: Faculty of Medicine. Total number of authors: 1 General rights Unless other specific re-use rights are stated the following general rights apply: Copyright and moral rights for the publications made accessible in the public portal are retained by the authors and/or other copyright owners and it is a condition of accessing publications that users recognise and abide by the legal requirements associated with these rights. • Users may download and print one copy of any publication from the public portal for the purpose of private study or research. • You may not further distribute the material or use it for any profit-making activity or commercial gain • You may freely distribute the URL identifying the publication in the public portal Read more about Creative commons licenses: https://creativecommons.org/licenses/ Take down policy If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim. LUND UNIVERSITY PO Box 117 221 00 Lund +46 46-222 00 00 Download date: 07. Oct. 2021 Division of Hematology and Transfusion Medicine Lund University, Lund, Sweden WILMS’ TUMOUR GENE 1 PROTEIN (WT1) – AN EFFECTOR IN LEUKEMOGENESIS? Karina Vidovic Thesis 2010 Contact adress Karina Vidovic Division of Hematology and Transfusion Medicine BMC, C14 Klinikgatan 28 SE- 221 84 Lund Sweden Phone +46 46 222 07 30 e-mail: [email protected] ISBN 978-91-86443-99-3 ! Karina Vidovic Printed by Media-Tryck, Lund, Sweden 2 ”The journey of a thousand miles begins with one step” Lao Tzu (Chinese taoist), 600-531 BC 3 4 LIST OF PAPERS This thesis is based on the following papers, referred to in the text by their Roman numerals I. -
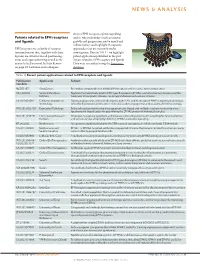
Patents Related to EPH Receptors and Ligands
NEWS & ANALYSIS discuss EPH receptor–ephrin signalling Patents related to EPH receptors and its role in disorders such as tumour and ligands growth and progression, nerve injury and inflammation, and highlight therapeutic EPH receptors are a family of receptor approaches that are currently under tyrosine kinases that, together with their investigation. Here in TABLE 1 we highlight ligands, are involved in cell positioning, patent applications published in the past tissue and organ patterning as well as the 3 years related to EPH receptors and ligands. control of cell survival. In their Review Data were researched using the Espacenet on page 39, Lackman and colleagues database. Table 1 | Recent patent applications related to EPH receptors and ligands Nature Reviews | Drug Discovery Publication Applicants Subject numbers NZ 581397 AstraZeneca Pyrimidine compounds that inhibit EPH receptors and are useful for treating cancer HK 1108702 Sanford-Burnham Peptides that selectively bind to EPH type-B receptors (EPHBs); useful for tumour imaging and the Institute treatment of neoplastic disease, neurological disease and vascular disease US 2013091591 California Institute of During angiogenesis, arterial cells express ephrin B2, and its receptor EPHB4 is expressed on venous Technology cells; this distinction can be used in methods to alter angiogenesis and to assess the effect of drugs WO 2013052710 Expression Pathology Selected reaction monitoring mass spectrometry-based and multiple reaction monitoring mass spectrometry-based assays for quantifying -

Transcriptional Regulation of RKIP in Prostate Cancer Progression
Health Science Campus FINAL APPROVAL OF DISSERTATION Doctor of Philosophy in Biomedical Sciences Transcriptional Regulation of RKIP in Prostate Cancer Progression Submitted by: Sandra Marie Beach In partial fulfillment of the requirements for the degree of Doctor of Philosophy in Biomedical Sciences Examination Committee Major Advisor: Kam Yeung, Ph.D. Academic William Maltese, Ph.D. Advisory Committee: Sonia Najjar, Ph.D. Han-Fei Ding, M.D., Ph.D. Manohar Ratnam, Ph.D. Senior Associate Dean College of Graduate Studies Michael S. Bisesi, Ph.D. Date of Defense: May 16, 2007 Transcriptional Regulation of RKIP in Prostate Cancer Progression Sandra Beach University of Toledo ACKNOWLDEGMENTS I thank my major advisor, Dr. Kam Yeung, for the opportunity to pursue my degree in his laboratory. I am also indebted to my advisory committee members past and present, Drs. Sonia Najjar, Han-Fei Ding, Manohar Ratnam, James Trempe, and Douglas Pittman for generously and judiciously guiding my studies and sharing reagents and equipment. I owe extended thanks to Dr. William Maltese as a committee member and chairman of my department for supporting my degree progress. The entire Department of Biochemistry and Cancer Biology has been most kind and helpful to me. Drs. Roy Collaco and Hong-Juan Cui have shared their excellent technical and practical advice with me throughout my studies. I thank members of the Yeung laboratory, Dr. Sungdae Park, Hui Hui Tang, Miranda Yeung for their support and collegiality. The data mining studies herein would not have been possible without the helpful advice of Dr. Robert Trumbly. I am also grateful for the exceptional assistance and shared microarray data of Dr. -

942.Full.Pdf
Original Article Opposite Effect of JAK2 on Insulin-Dependent Activation of Mitogen-Activated Protein Kinases and Akt in Muscle Cells Possible Target to Ameliorate Insulin Resistance Ana C.P. Thirone, Lellean JeBailey, Philip J. Bilan, and Amira Klip Many cytokines increase their receptor affinity for Janus kinases (JAKs). Activated JAK binds to signal transducers and activators of transcription, insulin receptor substrates olypeptides such as erythropoietin, prolactin, (IRSs), and Shc. Intriguingly, insulin acting through its leptin, angiotensin, growth hormone, most inter- own receptor kinase also activates JAK2. However, the leukins, and interferon-␥ bind to receptors that impact of such activation on insulin action remains un- Plack intrinsic kinase activity, recruiting and acti- known. To determine the contribution of JAK2 to insulin vating cytoplasmic tyrosine kinases of the Janus family signaling, we transfected L6 myotubes with siRNA against (JAK) consisting of JAK1, JAK2, JAK3, and Tyk2 (1–3). JAK2 (siJAK2), reducing JAK2 protein expression by 75%. Activated JAK phosphorylates tyrosine residues within Insulin-dependent phosphorylation of IRS1/2 and Shc was not affected by siJAK2, but insulin-induced phosphoryla- itself and the associated receptor forming high-affinity tion of the mitogen-activated protein kinases (MAPKs) binding sites for a variety of signaling proteins containing Src homology 2 and other phosphotyrosine-binding do- extracellular signal–related kinase, p38, and Jun NH2- terminal kinase and their respective upstream kinases mains, including signal transducers and activators of tran- MKK1/2, MKK3/6, and MKK4/7 was significantly lowered scription, insulin receptor substrates (IRSs), and the when JAK2 was depleted, correlating with a significant adaptor protein Shc (1–4). -

JAK Inhibitors for Treatment of Psoriasis: Focus on Selective TYK2 Inhibitors
Drugs https://doi.org/10.1007/s40265-020-01261-8 CURRENT OPINION JAK Inhibitors for Treatment of Psoriasis: Focus on Selective TYK2 Inhibitors Miguel Nogueira1 · Luis Puig2 · Tiago Torres1,3 © Springer Nature Switzerland AG 2020 Abstract Despite advances in the treatment of psoriasis, there is an unmet need for efective and safe oral treatments. The Janus Kinase– Signal Transducer and Activator of Transcription (JAK–STAT) pathway plays a signifcant role in intracellular signalling of cytokines of numerous cellular processes, important in both normal and pathological states of immune-mediated infamma- tory diseases. Particularly in psoriasis, where the interleukin (IL)-23/IL-17 axis is currently considered the crucial pathogenic pathway, blocking the JAK–STAT pathway with small molecules would be expected to be clinically efective. However, relative non-specifcity and low therapeutic index of the available JAK inhibitors have delayed their integration into the therapeutic armamentarium of psoriasis. Current research appears to be focused on Tyrosine kinase 2 (TYK2), the frst described member of the JAK family. Data from the Phase II trial of BMS-986165—a selective TYK2 inhibitor—in psoriasis have been published and clinical results are encouraging, with a large Phase III programme ongoing. Further, the selective TYK2 inhibitor PF-06826647 is being tested in moderate-to-severe psoriasis in a Phase II clinical trial. Brepocitinib, a potent TYK2/JAK1 inhibitor, is also being evaluated, as both oral and topical treatment. Results of studies with TYK2 inhibitors will be important in assessing the clinical efcacy and safety of these drugs and their place in the therapeutic armamentarium of psoriasis. -

Ephb3 Suppresses Non-Small-Cell Lung Cancer Metastasis Via a PP2A/RACK1/Akt Signalling Complex
ARTICLE Received 7 Nov 2011 | Accepted 11 Jan 2012 | Published 7 Feb 2012 DOI: 10.1038/ncomms1675 EphB3 suppresses non-small-cell lung cancer metastasis via a PP2A/RACK1/Akt signalling complex Guo Li1, Xiao-Dan Ji1, Hong Gao1, Jiang-Sha Zhao1, Jun-Feng Xu1, Zhi-Jian Sun1, Yue-Zhen Deng1, Shuo Shi1, Yu-Xiong Feng1, Yin-Qiu Zhu1, Tao Wang2, Jing-Jing Li1 & Dong Xie1 Eph receptors are implicated in regulating the malignant progression of cancer. Here we find that despite overexpression of EphB3 in human non-small-cell lung cancer, as reported previously, the expression of its cognate ligands, either ephrin-B1 or ephrin-B2, is significantly downregulated, leading to reduced tyrosine phosphorylation of EphB3. Forced activation of EphB3 kinase in EphB3-overexpressing non-small-cell lung cancer cells inhibits cell migratory capability in vitro as well as metastatic seeding in vivo. Furthermore, we identify a novel EphB3-binding protein, the receptor for activated C-kinase 1, which mediates the assembly of a ternary signal complex comprising protein phosphatase 2A, Akt and itself in response to EphB3 activation, leading to reduced Akt phosphorylation and subsequent inhibition of cell migration. Our study reveals a novel tumour-suppressive signalling pathway associated with kinase-activated EphB3 in non-small-cell lung cancer, and provides a potential therapeutic strategy by activating EphB3 signalling, thus inhibiting tumour metastasis. 1 Key Laboratory of Nutrition and Metabolism, Institute for Nutritional Sciences, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences and Graduate School of Chinese Academy of Sciences, Shanghai 200031, China. 2 The Eastern Hepatobiliary Surgery Hospital, the Second Military Medical University, Shanghai 200433, China. -

Original Article ERBB3, IGF1R, and TGFBR2 Expression Correlate With
Am J Cancer Res 2018;8(5):792-809 www.ajcr.us /ISSN:2156-6976/ajcr0077452 Original Article ERBB3, IGF1R, and TGFBR2 expression correlate with PDGFR expression in glioblastoma and participate in PDGFR inhibitor resistance of glioblastoma cells Kang Song1,2*, Ye Yuan1,2*, Yong Lin1,2, Yan-Xia Wang1,2, Jie Zhou1,2, Qu-Jing Gai1,2, Lin Zhang1,2, Min Mao1,2, Xiao-Xue Yao1,2, Yan Qin1,2, Hui-Min Lu1,2, Xiang Zhang1,2, You-Hong Cui1,2, Xiu-Wu Bian1,2, Xia Zhang1,2, Yan Wang1,2 1Department of Pathology, Institute of Pathology and Southwest Cancer Center, Southwest Hospital, Third Military Medical University, Chongqing 400038, China; 2Key Laboratory of Tumor Immunology and Pathology of Ministry of Education, Chongqing 400038, China. *Equal contributors. Received April 6, 2018; Accepted April 9, 2018; Epub May 1, 2018; Published May 15, 2018 Abstract: Glioma, the most prevalent malignancy in brain, is classified into four grades (I, II, III, and IV), and grade IV glioma is also known as glioblastoma multiforme (GBM). Aberrant activation of receptor tyrosine kinases (RTKs), including platelet-derived growth factor receptor (PDGFR), are frequently observed in glioma. Accumulating evi- dence suggests that PDGFR plays critical roles during glioma development and progression and is a promising drug target for GBM therapy. However, PDGFR inhibitor (PDGFRi) has failed in clinical trials, at least partially, due to the activation of other RTKs, which compensates for PDGFR inhibition and renders tumor cells resistance to PDGFRi. Therefore, identifying the RTKs responsible for PDGFRi resistance might provide new therapeutic targets to syner- getically enhance the efficacy of PDGFRi. -

Supplementary Table 1: Adhesion Genes Data Set
Supplementary Table 1: Adhesion genes data set PROBE Entrez Gene ID Celera Gene ID Gene_Symbol Gene_Name 160832 1 hCG201364.3 A1BG alpha-1-B glycoprotein 223658 1 hCG201364.3 A1BG alpha-1-B glycoprotein 212988 102 hCG40040.3 ADAM10 ADAM metallopeptidase domain 10 133411 4185 hCG28232.2 ADAM11 ADAM metallopeptidase domain 11 110695 8038 hCG40937.4 ADAM12 ADAM metallopeptidase domain 12 (meltrin alpha) 195222 8038 hCG40937.4 ADAM12 ADAM metallopeptidase domain 12 (meltrin alpha) 165344 8751 hCG20021.3 ADAM15 ADAM metallopeptidase domain 15 (metargidin) 189065 6868 null ADAM17 ADAM metallopeptidase domain 17 (tumor necrosis factor, alpha, converting enzyme) 108119 8728 hCG15398.4 ADAM19 ADAM metallopeptidase domain 19 (meltrin beta) 117763 8748 hCG20675.3 ADAM20 ADAM metallopeptidase domain 20 126448 8747 hCG1785634.2 ADAM21 ADAM metallopeptidase domain 21 208981 8747 hCG1785634.2|hCG2042897 ADAM21 ADAM metallopeptidase domain 21 180903 53616 hCG17212.4 ADAM22 ADAM metallopeptidase domain 22 177272 8745 hCG1811623.1 ADAM23 ADAM metallopeptidase domain 23 102384 10863 hCG1818505.1 ADAM28 ADAM metallopeptidase domain 28 119968 11086 hCG1786734.2 ADAM29 ADAM metallopeptidase domain 29 205542 11085 hCG1997196.1 ADAM30 ADAM metallopeptidase domain 30 148417 80332 hCG39255.4 ADAM33 ADAM metallopeptidase domain 33 140492 8756 hCG1789002.2 ADAM7 ADAM metallopeptidase domain 7 122603 101 hCG1816947.1 ADAM8 ADAM metallopeptidase domain 8 183965 8754 hCG1996391 ADAM9 ADAM metallopeptidase domain 9 (meltrin gamma) 129974 27299 hCG15447.3 ADAMDEC1 ADAM-like,