Durham E-Theses
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

2019 Cruise Directory
Despite the modern fashion for large floating resorts, we b 7 nights 0 2019 CRUISE DIRECTORY Highlands and Islands of Scotland Orkney and Shetland Northern Ireland and The Isle of Man Cape Wrath Scrabster SCOTLAND Kinlochbervie Wick and IRELAND HANDA ISLAND Loch a’ FLANNAN Stornoway Chàirn Bhain ISLES LEWIS Lochinver SUMMER ISLES NORTH SHIANT ISLES ST KILDA Tarbert SEA Ullapool HARRIS Loch Ewe Loch Broom BERNERAY Trotternish Inverewe ATLANTIC NORTH Peninsula Inner Gairloch OCEAN UIST North INVERGORDON Minch Sound Lochmaddy Uig Shieldaig BENBECULA Dunvegan RAASAY INVERNESS SKYE Portree Loch Carron Loch Harport Kyle of Plockton SOUTH Lochalsh UIST Lochboisdale Loch Coruisk Little Minch Loch Hourn ERISKAY CANNA Armadale BARRA RUM Inverie Castlebay Sound of VATERSAY Sleat SCOTLAND PABBAY EIGG MINGULAY MUCK Fort William BARRA HEAD Sea of the Glenmore Loch Linnhe Hebrides Kilchoan Bay Salen CARNA Ballachulish COLL Sound Loch Sunart Tobermory Loch à Choire TIREE ULVA of Mull MULL ISLE OF ERISKA LUNGA Craignure Dunsta!nage STAFFA OBAN IONA KERRERA Firth of Lorn Craobh Haven Inveraray Ardfern Strachur Crarae Loch Goil COLONSAY Crinan Loch Loch Long Tayvallich Rhu LochStriven Fyne Holy Loch JURA GREENOCK Loch na Mile Tarbert Portavadie GLASGOW ISLAY Rothesay BUTE Largs GIGHA GREAT CUMBRAE Port Ellen Lochranza LITTLE CUMBRAE Brodick HOLY Troon ISLE ARRAN Campbeltown Firth of Clyde RATHLIN ISLAND SANDA ISLAND AILSA Ballycastle CRAIG North Channel NORTHERN Larne IRELAND Bangor ENGLAND BELFAST Strangford Lough IRISH SEA ISLE OF MAN EIRE Peel Douglas ORKNEY and Muckle Flugga UNST SHETLAND Baltasound YELL Burravoe Lunna Voe WHALSAY SHETLAND Lerwick Scalloway BRESSAY Grutness FAIR ISLE ATLANTIC OCEAN WESTRAY SANDAY STRONSAY ORKNEY Kirkwall Stromness Scapa Flow HOY Lyness SOUTH RONALDSAY NORTH SEA Pentland Firth STROMA Scrabster Caithness Wick Welcome to the 2019 Hebridean Princess Cruise Directory Unlike most cruise companies, Hebridean operates just one very small and special ship – Hebridean Princess. -

Endurance Door Brochure
PROTECT YOUR HOME WITH A endurancedoors.co.uk SOLID AND SECURE DOOR INTRODUCTION 1 Safety, Security & Style. It starts with an CONTENTS Endurance door. Why Endurance? 4-5 Endurance Doors are renowned for their strength and security, without compromising on style. DNA of an Endurance Door 6 Secured by Design 7 At 48mm thick and with cross-bonded laminations, an Endurance door delivers unrivaled strength and Preferred Installer Network 8 dimensional stability. It’s the frst choice in delivering the Design Your Dream Door 9 highest level of security, providing home owners with peace of mind. Colours 10-11 Classic Collection 12-37 Made up of 17 laminations, Endurance Doors are almost 10% thicker than most composite doors, guaranteeing Country Collection 38-55 the safety and security of your family and your home. Urban Collection 56-69 With over 50 diferent door designs to choose from, Other Door Styles available in a wide selection of colours with an array & Accessories 70- 85 of diferent door furniture and glazing options, your Endurance Door can be as individual as you are. Glazing Styles 86-87 Glass Matrix 88-89 The Green Promise 90 Quality Standards 91 Classic Collection Country Collection Urban Collection Pages: 12-37 Pages: 38-55 Pages: 56-69 2 VISIT ENDURANCEDOORS.CO.UK TO DESIGN YOUR DOOR INTRODUCTION 3 WHY ENDURANCE? Endurance Doors are renowned for Detailed embossed wood grain texture door their strength and Secured by Design skins, traditional look security, without Secured By Design is the ofcial UK Police with modern technology fagship initiative supporting the principles of ‘designing out crime’. -

Is Wales' Highest Mountain the Perfect Starter Peak for Kids?
SNOWDON FOR ALL CHILD’S PLAY Is Wales’ highest mountain the perfect starter peak for kids? We sent a rock star to find out... WORDS & PHOTOGRAPHS PHOEBE SMITH ver half a million an ideal first mountain for kids visitors a year would to climb. Naturally, we wanted to suggest the cat is well put that theory to the test, so we and truly out of the went in search of an adventurous bag with Snowdon. family looking for their first taste Arguably, it’s the perfect of proper mountain walking. We mountain for walkers. weren’t expecting that search to Undeniably, it’s one of lead us to a BBC radio presenter OEurope’s most spectacular. This is who also happened to be the lead a peak of extraordinary, unrivalled singer of a multi-million-selling versatility, one that’s historically 1990s rock band. But that’s been used as a training ground exactly what happened. for Everest-bound mountaineers, The message arrived quite but also one where you could unexpectedly one Wednesday achievably stroll with your afternoon. Scanning through my children to the summit. emails, it was a pretty normal day. Then I saw it, the There are no fewer than 10 recognised ways one that stood out above the rest. The subject line to walk or scramble to Snowdon’s pyramidal read: ‘SNOWDONIA – February half-term?’ 1085m top. The beginner-friendly Llanberis Path The message was from Cerys Matthews, the offers the most pedestrian ascent; the South Ridge former frontwoman of rock band Catatonia and holds the key to the mountain’s secret back door; a current BBC Radio 6 Music presenter, who I’d while the notoriously nerve-zapping and razor-sharp accompanied on a wild camping trip a few months ridgeline of Crib Goch is reserved for those with a earlier. -

NLCA06 Snowdonia - Page 1 of 12
National Landscape Character 31/03/2014 NLCA06 Snowdonia Eryri – Disgrifiad cryno Dyma fro eang, wledig, uchel, sy’n cyd-ffinio’n fras â Pharc Cenedlaethol Eryri. Ei nodwedd bennaf yw ei mynyddoedd, o ba rai yr Wyddfa yw mynydd uchaf Cymru a Lloegr, yn 3560’ (1085m) o uchder. Mae’r mynyddoedd eraill yn cynnwys y Carneddau a’r Glyderau yn y gogledd, a’r Rhinogydd a Chadair Idris yn y de. Yma ceir llawer o fryndir mwyaf trawiadol y wlad, gan gynnwys pob un o gopaon Cymru sy’n uwch na 3,000 o droedfeddi. Mae llawer o nodweddion rhewlifol, gan gynnwys cribau llymion, cymoedd, clogwyni, llynnoedd (gan gynnwys Llyn Tegid, llyn mwyaf Cymru), corsydd, afonydd a rhaeadrau. Mae natur serth y tir yn gwneud teithio’n anodd, a chyfyngir mwyafrif y prif ffyrdd i waelodion dyffrynnoedd a thros fylchau uchel. Yn ddaearegol, mae’n ardal amrywiol, a fu â rhan bwysig yn natblygiad cynnar gwyddor daeareg. Denodd sylw rhai o sylfaenwyr yr wyddor, gan gynnwys Charles Darwin, a archwiliodd yr ardal ym 1831. Y mae ymhell, fodd bynnag, o fod yn ddim ond anialdir uchel. Am ganrifoedd, bu’r ardal yn arwydd ysbryd a rhyddid y wlad a’i phobl. Sefydlwyd bwrdeistrefi Dolgellau a’r Bala yng nghyfnod annibyniaeth Cymru cyn y goresgyniad Eingl-normanaidd. Felly, hefyd, llawer o aneddiadau llai ond hynafol fel Dinas Mawddwy. O’i ganolfan yn y Bala, dechreuodd y diwygiad Methodistaidd ar waith trawsffurfio Cymru a’r ffordd Gymreig o fyw yn y 18fed ganrif a’r 19eg. Y Gymraeg yw iaith mwyafrif y trigolion heddiw. -

GUIDE for GROUPS an Ancient Norman Family, the Hansards, Who Arrived in England with William the 0844 387 6680 Conqueror
ULTIMATE STATUS SYMBOL There’s more to explore with Best Western Best Western Walworth Castle Hotel near Darlington looks like a real castle but its imposing turrets and arrow slits are designed for decoration not defence. In 1600, when the extraordinary house was being built for Thomas Jenison, all the best families had castles and so the features were added to give it a fashionably grand appearance. ‘As Visit: far as we know, it was never used as a defensive fort,’ says the hotel manager. www.bestwestern.co.uk/groups 2019 However, a true castle did stand on the site in the 12th century; it was the home of Call us on: GUIDE FOR GROUPS an ancient Norman family, the Hansards, who arrived in England with William the 0844 387 6680 Conqueror. Calling from overseas: +44 1904 695450 Email: [email protected] bestwestern.co.uk Front cover images: Llandwyn Island, Wales | York Minster, York 2019 Loch Lomand, Scotland | Best Western Walton Park Hotel, Clevedon TO TELL TO MANY ASTORY heavily in the box offi cesmash,HotFuzz. offi inthebox heavily onfi hasbeencaptured hotel the recently, More localstations. three from pickuppassengers to conveyance itsown ran whenthehotel of theTrain theAge Anne ofDenmarkandwitnessed in honour of afeast hosted the citizens, theBishopsand between the disputes hasexperienced Wells Hotel, Plus Swan Western theBest thecenturies Over InnsofWells. onThe Scrase Tony by mentionedinanarticle1422, whenitwas atleast backto date parts ofthehotel oldest The people. famous many to host houseandhasplayed a majorposting once -
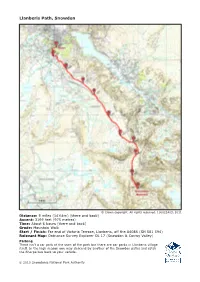
Snpa-Llanberis-Path-Map.Pdf
Llanberis Path, Snowdon © Crown copyright. All rights reserved. 100022403, 2011 Distance: 9 miles (14½km) (there and back) Ascent: 3199 feet (975 metres) Time: About 6 hours (there and back) Grade: Mountain Walk Start / Finish: Far end of Victoria Terrace, Llanberis, off the A4086 (SH 581 594) Relevant Map: Ordnance Survey Explorer OL 17 (Snowdon & Conwy Valley) Parking There isn’t a car park at the start of the path but there are car parks in Llanberis village itself. In the high season you may descend by another of the Snowdon paths and catch the Sherpa bus back to your vehicle. © 2010 Snowdonia National Park Authority Llanberis Path, Snowdon Llanberis Path is the longest and most gradual of the six main paths to the summit of Snowdon, and offers fantastic views of Cwm Brwynog, Llanberis and over the Menai straights towards Anglesey. This is the most popular path amongst leisurely walkers as it is thought to be the easiest to walk in mild weather, but in winter, the highest slopes of the path can become very dangerous. The path mainly follows the Snowdon Mountain Railway track, and goes by Hebron, Halfway and Clogwyn stations. Before the railway was opened in 1896, visitors employed guides to lead them to the summit along this path on mule-back. A remarkable geological feature can be seen from the Llanberis path, the Clogwyn Du’r Arddu syncline. The syncline was formed over thousands of years, as the earth moved and transformed horizontal depos- its into vertical layers of rock. Safety Note The Llanberis Path and Snowdon Mountain Railway cross above Clogwyn Coch. -

Welsh 3 Peaks Challenge
WELSH 3 PEAKS CHALLENGE Highlights • 3 peaks! 2334 metres of ascent! 17 miles trekked! In 1-day! Tick! • One of the classic walking challenges in Wales with summiting the three highest peaks • Conquer Snowdon, Cader Idris and Pen-y-Fan • Ascend ridges, cross linked peaks and experience the stellar views and natural beauty of Snowdonia, home to the highest peak in Wales • Trek with pride knowing you are helping support the work of the Snowdonia Society and Cool Earth • Accompanied by local Adventurous Ewe Mountain Leaders • New safety and wellbeing guidelines are in place for this adventure • We look forward to welcoming you to our beautiful homeland of Wales. • #ewecandoit www.yourcompany.com 1 WELCOME Overview Are you ready for an epic mountain adventure embracing the rugged mountains of Wales? Conquering the three highest peaks in Wales, this journey will take you through spectacular landscapes and test your mettle on a tough 1-day adventure. The Welsh 3 Peaks Challenge is made up of three of the highest and most iconic mountains in Wales: Snowdon, Wales’ tallest peak and the highest point in Britain outside the Scottish highlands; Cader Idris, a spectacular peak at the southerly edge of Snowdonia National Park; and Pen y Fan, the highest peak in the Brecon Beacons National Park in South Wales. There’s plenty of mythical legends surrounding these mountains and your local leaders will keep you entertained (or pre-occupied) with stories of giants, villians and poets or explain some of the spectacular geology before you’re greeted with 360 views from each mountain summit, weather permitting of course. -

Required Equipment - Kit Checklist
Required Equipment - Kit Checklist The following items must be carried on all mountains by each team. Each team will be checked for all these items during registration. Subsequent checks will be made before each mountain stage of the event. Team equipment: ¨ First-aid kit (remember special needs of team members i.e. asthmatic etc.) ¨ Compass ¨ Maps (Snowdon, Cadair Idris, Pen-y-Fan) The maps you will require for each mountain are: Snowdon: Ordnance Survey Explorer OL No 17 (1 to 25,000) “Snowdon and Conwy Valley” Cadair Idris: Ordnance Survey Explorer OL No 23 (1 to 25,000) “Cadair Idris and Bala Lake” Pen y Fan: Ordnance Survey Explorer OL No 12 (1 to 25,000) “Brecon Beacons National Park – Western and Central areas” (see maps section of fundraising pack for helpful information) ¨ Note pad and pencil ¨ Mobile phone þ Bothie (will be issued at team briefing) þ Mountain Passport (will be issued at team briefing) Individual requirements: ¨ Rucksack (approx. 30-40 litres) ¨ Waterproof liner ¨ Appropriate footwear (see details below) ¨ Survival bag A survival bag is a person-sized waterproof bag, typically orange in colour, designed to avert the threat of hypothermia from exposure. It is reasonably light, made from strong, waterproof and tear-proof plastic, and provides some amount of thermal insulation and can be purchased at most outdoor stores and online for less than £5. ¨ Set of waterproofs (jacket & trousers) ¨ Hat and gloves ¨ Whistle ¨ Emergency rations (chocolate, dried fruit, nuts, cereal bars etc.) ¨ Torch ¨ Money (in case of emergency) ¨ Drink The amount of fluid required per person will change depending on the weather conditions. -

Hill Walking & Mountaineering
Hill Walking & Mountaineering in Snowdonia Introduction The craggy heights of Snowdonia are justly regarded as the finest mountain range south of the Scottish Highlands. There is a different appeal to Snowdonia than, within the picturesque hills of, say, Cumbria, where cosy woodland seems to nestle in every valley and each hillside seems neatly manicured. Snowdonia’s hillsides are often rock strewn with deep rugged cwms biting into the flank of virtually every mountainside, sometimes converging from two directions to form soaring ridges which lead to lofty peaks. The proximity of the sea ensures that a fine day affords wonderful views, equally divided between the ever- changing seas and the serried ranks of mountains fading away into the distance. Eryri is the correct Welsh version of the area the English call Snowdonia; Yr Wyddfa is similarly the correct name for the summit of Snowdon, although Snowdon is often used to demarcate the whole massif around the summit. The mountains of Snowdonia stretch nearly fifty miles from the northern heights of the Carneddau, looming darkly over Conwy Bay, to the southern fringes of the Cadair Idris massif, overlooking the tranquil estuary of the Afon Dyfi and Cardigan Bay. From the western end of the Nantlle Ridge to the eastern borders of the Aran range is around twenty- five miles. Within this area lie nine distinct mountain groups containing a wealth of mountain walking possibilities, while just outside the National Park, the Rivals sit astride the Lleyn Peninsula and the Berwyns roll upwards to the east of Bala. The traditional bases of Llanberis, Bethesda, Capel Curig, Betws y Coed and Beddgelert serve the northern hills and in the south Barmouth, Dinas Mawddwy, Dolgellau, Tywyn, Machynlleth and Bala provide good locations for accessing the mountains. -

Nor' West News
Nor’ West News The NeWSletter of the Nor’ West Sgurramblers Issue No. 144 – October 2010 www.sgurramblers.org.uk MEET REPORTS The Joys of Scottish Summer Weather : Meet No. 271 (Friday 9 – Sunday 18 July 2010) Roger Reeves and Brian Billington came to pick me up on the first Friday after spending Thursday night at Alston youth hostel in Northumberland. And I was grateful for their help in packing a mountain of food into Roger’s car. I had been concerned that Roger might not have sufficient room for all my goods but, to my relief, he demonstrated that he has got packing down to a fine art. We arrived at the “My Highland Bunkhouse” near Newtonmore in the late afternoon. Hugh Toal, our guest on the first part of this NWS meet, arrived shortly after us. The bunkhouse proved to be a very modernistic structure, built the previous year adjacent to the equally modernistic and aptly named “Round House” farmhouse (also of very recent construction). Roger remarked that the buildings were like something out of “Harry Potter” but, thankfully, he stopped short of extending the analogy from the buildings to those who attended either part of this two part meet. A long winding track led from the road to the bunkhouse which only sleeps seven. Five of us had booked for this part of the meet: so we had it for our exclusive use. At £70 a night it was good value. The bunkhouse was also in a lovely location off the Newtonmore/Laggan road, though not the easiest place to find. -

Cicerone-Catalogue.Pdf
SPRING/SUMMER CATALOGUE 2020 Cover: A steep climb to Marions Peak from Hiking the Overland Track by Warwick Sprawson Photo: ‘The veranda at New Pelion Hut – attractive habitat for shoes and socks’ also from Hiking the Overland Track by Warwick Sprawson 2 | BookSource orders: tel 0845 370 0067 [email protected] Welcome to CICERONE Nearly 400 practical and inspirational guidebooks for hikers, mountaineers, climbers, runners and cyclists Contents The essence of Cicerone ..................4 Austria .................................38 Cicerone guides – unique and special ......5 Eastern Europe ..........................38 Series overview ........................ 6-9 France, Belgium, Luxembourg ............39 Spotlight on new titles Spring 2020 . .10–21 Germany ...............................41 New title summary January – June 2020 . .21 Ireland .................................41 Italy ....................................42 Mediterranean ..........................43 Book listing New Zealand and Australia ...............44 North America ..........................44 British Isles Challenges, South America ..........................44 Collections and Activities ................22 Scandinavia, Iceland and Greenland .......44 Scotland ................................23 Slovenia, Croatia, Montenegro, Albania ....45 Northern England Trails ..................26 Spain and Portugal ......................45 North East England, Yorkshire Dales Switzerland .............................48 and Pennines ...........................27 Japan, Asia -

North West England & Isle of Man: Climate
North West England & Isle of Man: climate This describes the main features of the climate of Cheshire, Merseyside, Greater Manchester, Lancashire, Cumbria, part of North Yorkshire and the Isle of Man. The eastern boundary of the region lies along the watershed of the Pennines. There are several distinct geographical areas within the region. Much of the area west of the Pennines from Lancashire southwards forms part of a plain, mainly below 150 metres, with westward draining rivers such as the Mersey, Ribble and Lune. Liverpool and Manchester are the main centres of population. To the east, the Pennines are a chain of rolling gritstone moors rising to well over 600 metres and reaching their highest point at Cross Fell (893 metres). The Pennines form a natural barrier to east-west communications, but there are the Tyne gap linking Carlisle and Newcastle and the Aire gap linking Lancashire and Yorkshire. The Lake District in Cumbria includes Scafell Pike (978 metres), the highest mountain in England, and Windermere, the largest lake. To the north and NE, there is further low ground (below 60 metres) in the Vale of Eden and the Solway Plain, close to Carlisle. Most of the Isle of Man consists of a central mountain mass rising to 621 metres on Snaefell. Douglas is the main town and the centre of the island's tourist industry. The range of topography and altitude found in both NW England and the Isle of Man provide a climate of great variety, and the region includes both the coldest place in England (Cross Fell in the Pennines) and the wettest place in England (the Lakeland fells around Seathwaite in Cumbria).