Heterogeneity of Human Lactoferrin Due to Differences in Sialic Acid Content
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Types of Acute Phase Reactants and Their Importance in Vaccination (Review)
BIOMEDICAL REPORTS 12: 143-152, 2020 Types of acute phase reactants and their importance in vaccination (Review) RAFAAT H. KHALIL1 and NABIL AL-HUMADI2 1Department of Biology, College of Science and Technology, Florida Agricultural and Mechanical University, Tallahassee, FL 32307; 2Office of Vaccines, Food and Drug Administration, Center for Biologics Evaluation and Research, Silver Spring, MD 20993, USA Received May 10, 2019; Accepted November 25, 2019 DOI: 10.3892/br.2020.1276 Abstract. Vaccines are considered to be one of the most human and veterinary medicine. Proteins which are expressed cost-effective life-saving interventions in human history. in the acute phase are potential biomarkers for the diagnosis The body's inflammatory response to vaccines has both of inflammatory disease, for example, acute phase proteins desired effects (immune response), undesired effects [(acute (APPs) are indicators of successful organ transplantation phase reactions (APRs)] and trade‑offs. Trade‑offs are and can be used to predict the ameliorative effect of cancer more potent immune responses which may be potentially therapy (1,2). APPs are primarily synthesized in hepatocytes. difficult to separate from potent acute phase reactions. The acute phase response is a spontaneous reaction triggered Thus, studying acute phase proteins (APPs) during vaccina- by disrupted homeostasis resulting from environmental distur- tion may aid our understanding of APRs and homeostatic bances (3). Acute phase reactions (APRs) usually stabilize changes which can result from inflammatory responses. quickly, after recovering from a disruption to homeostasis Depending on the severity of the response in humans, these within a few days to weeks; however, APPs expression levels reactions can be classified as major, moderate or minor. -

Lactoferrin and Its Detection Methods: a Review
nutrients Review Lactoferrin and Its Detection Methods: A Review Yingqi Zhang, Chao Lu and Jin Zhang * Department of Chemical and Biochemical Engineering, University of Western Ontario, London, ON N6A 5B9, Canada; [email protected] (Y.Z.); [email protected] (C.L.) * Correspondence: [email protected] Abstract: Lactoferrin (LF) is one of the major functional proteins in maintaining human health due to its antioxidant, antibacterial, antiviral, and anti-inflammatory activities. Abnormal levels of LF in the human body are related to some serious diseases, such as inflammatory bowel disease, Alzheimer’s disease and dry eye disease. Recent studies indicate that LF can be used as a biomarker for diagnosis of these diseases. Many methods have been developed to detect the level of LF. In this review, the biofunctions of LF and its potential to work as a biomarker are introduced. In addition, the current methods of detecting lactoferrin have been presented and discussed. We hope that this review will inspire efforts in the development of new sensing systems for LF detection. Keywords: lactoferrin; biomarkers; immunoassay; instrumental analysis; sensor 1. Introduction Lactoferrin (known as lactotransferrin, LF), with a molecular weight of about 80 kDa, is a functional glycoprotein, which contains about 690 amino acid residues. It was first isolated from bovine milk by Sorensen in 1939 and was first isolated from human milk by Citation: Zhang, Y.; Lu, C.; Zhang, J. Johanson in 1960 [1,2]. The three-dimensional structure of LF has been unveiled by high Lactoferrin and Its Detection resolution X-ray crystallographic analysis, and it consists of two homologous globular lobes Methods: A Review. -

Influence of Infection and Inflammation on Biomarkers of Nutritional Status
A2.4 INFLUENCE OF INFECTION AND INFLAMMATION ON BIOMARKERS OF NUTRITIONAL STATUS A2.4 Influence of infection and inflammation on biomarkers of nutritional status with an emphasis on vitamin A and iron David I. Thurnham1 and George P. McCabe2 1 Northern Ireland Centre for Food and Health, University of Ulster, Coleraine, United Kingdom of Great Britain and Northern Ireland 2 Statistics Department, Purdue University, West Lafayette, Indiana, United States of America Corresponding author: David I. Thurnham; [email protected] Suggested citation: Thurnham DI, McCabe GP. Influence of infection and inflammation on biomarkers of nutritional status with an emphasis on vitamin A and iron. In: World Health Organization. Report: Priorities in the assessment of vitamin A and iron status in populations, Panama City, Panama, 15–17 September 2010. Geneva, World Health Organization, 2012. Abstract n Many plasma nutrients are influenced by infection or tissue damage. These effects may be passive and the result of changes in blood volume and capillary permeability. They may also be the direct effect of metabolic alterations that depress or increase the concentration of a nutrient or metabolite in the plasma. Where the nutrient or metabolite is a nutritional biomarker as in the case of plasma retinol, a depression in retinol concentrations will result in an overestimate of vitamin A deficiency. In contrast, where the biomarker is increased due to infection as in the case of plasma ferritin concentrations, inflammation will result in an underestimate of iron deficiency. Infection and tissue damage can be recognized by their clinical effects on the body but, unfortunately, subclinical infection or inflammation can only be recognized by measur- ing inflammation biomarkers in the blood. -

The Biology of Lactoferrin, an Iron-Binding Protein That Can Help Defend Against Viruses and Bacteria
Downloaded from orbit.dtu.dk on: Oct 02, 2021 The Biology of Lactoferrin, an Iron-Binding Protein That Can Help Defend Against Viruses and Bacteria Kell, Douglas B.; Heyden, Eugene L.; Pretorius, Etheresia Published in: Frontiers in Immunology Link to article, DOI: 10.3389/fimmu.2020.01221 Publication date: 2020 Document Version Publisher's PDF, also known as Version of record Link back to DTU Orbit Citation (APA): Kell, D. B., Heyden, E. L., & Pretorius, E. (2020). The Biology of Lactoferrin, an Iron-Binding Protein That Can Help Defend Against Viruses and Bacteria. Frontiers in Immunology, 11, [1221]. https://doi.org/10.3389/fimmu.2020.01221 General rights Copyright and moral rights for the publications made accessible in the public portal are retained by the authors and/or other copyright owners and it is a condition of accessing publications that users recognise and abide by the legal requirements associated with these rights. Users may download and print one copy of any publication from the public portal for the purpose of private study or research. You may not further distribute the material or use it for any profit-making activity or commercial gain You may freely distribute the URL identifying the publication in the public portal If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim. HYPOTHESIS AND THEORY published: 28 May 2020 doi: 10.3389/fimmu.2020.01221 The Biology of Lactoferrin, an Iron-Binding Protein That Can Help Defend Against Viruses and Bacteria Douglas B. -

Plasma Protein Binding Structure Activity Relationship Related to the N-Terminus of Daptomycin
Monash Institute of Pharmaceutical P1318 Sciences ECCMID 2017, Plasma protein binding structure activity relationship related to the N-terminus of daptomycin 381 Royal Parade, Parkville, VIC 3052 Melbourne, Australia Vienna 1 2 3 2 4 1 Tel: +61 3 9903 9539 Elena K. Schneider , Johnny X. Huang , Vincenzo Carbone , Matthew A. Cooper , Jian Li *, Tony Velkov * [email protected] 1Drug Development and Innovation, Drug Delivery, Disposition and Dynamics. Monash Institute of Pharmaceutical Sciences, Monash University, Australia. 2Institute for Molecular Bioscience, The University of Queensland St Lucia QLD 4072. 3Animal Nutrition and Health, Ag Research Limited, Grasslands Research Centre, Palmerston North, New Zealand. 4Monash Biomedicine Discovery Institute, Department of Microbiology, Monash University, Australia. Abstract Methods Results Background: Daptomycin is a lipopeptide antibiotic that is highly bound to plasma proteins. To date the plasma components and • Flourometric binding assay: • SPR: structure-activity relationships responsible for the plasma protein binding profile of daptomycin remain uncharacterized. We used the site selective fluorescent probes . Native daptomycin binds in order of decreasing Methods: dansylamide (site 1) and dansylsarcosine affinity to: HSA >> α-1-antitrypsin > low density (site 2) on HSA. As the control we employed lipoprotein (LDL) ≥ haemoglobin > sex hormone • In the present study we have employed surface plasmon resonance (SPR) and fluorescence displacement assay to Nifedipine for site 1 and ibuprofen -

Detailed Structure and Pathophysiological Roles of the Iga-Albumin Complex in Multiple Myeloma
International Journal of Molecular Sciences Article Detailed Structure and Pathophysiological Roles of the IgA-Albumin Complex in Multiple Myeloma Yuki Kawata 1, Hisashi Hirano 1, Ren Takahashi 1, Yukari Miyano 1, Ayuko Kimura 1, Natsumi Sato 1, Yukio Morita 2, Hirokazu Kimura 1,* and Kiyotaka Fujita 1 1 Department of Health Sciences, Gunma Paz University Graduate School of Health Sciences, 1-7-1, Tonyamachi, Takasaki-shi, Gunma 370-0006, Japan; [email protected] (Y.K.); [email protected] (H.H.); [email protected] (R.T.); [email protected] (Y.M.); [email protected] (A.K.); [email protected] (N.S.); [email protected] (K.F.) 2 Laboratory of Public Health II, Azabu University School of Veterinary Medicine, 1-17-71, Fuchinobe, Chuo-ku, Sagamihara, Kanagawa 252-5201, Japan; [email protected] * Correspondence: [email protected]; Tel.: +81-27-365-3366; Fax: +81-27-388-0386 Abstract: Immunoglobulin A (IgA)-albumin complexes may be associated with pathophysiology of multiple myeloma, although the etiology is not clear. Detailed structural analyses of these protein– protein complexes may contribute to our understanding of the pathophysiology of this disease. We analyzed the structure of the IgA-albumin complex using various electrophoresis, mass spectrom- etry, and in silico techniques. The data based on the electrophoresis and mass spectrometry showed that IgA in the sera of patients was dimeric, linked via the J chain. Only dimeric IgA can bind to albumin molecules leading to IgA-albumin complexes, although both monomeric and dimeric forms of IgA were present in the sera. -

Alpha -Antitrypsin Deficiency
The new england journal of medicine Review Article Dan L. Longo, M.D., Editor Alpha1-Antitrypsin Deficiency Pavel Strnad, M.D., Noel G. McElvaney, D.Sc., and David A. Lomas, Sc.D. lpha1-antitrypsin (AAT) deficiency is one of the most common From the Department of Internal Med genetic diseases. Most persons carry two copies of the wild-type M allele icine III, University Hospital RWTH of SERPINA1, which encodes AAT, and have normal circulating levels of the (Rheinisch–Westfälisch Technische Hoch A schule) Aachen, Aachen, Germany (P.S.); protein. Ninety-five percent of severe cases of AAT deficiency result from the homo- the Irish Centre for Genetic Lung Dis zygous substitution of a single amino acid, Glu342Lys (the Z allele), which is present ease, Royal College of Surgeons in Ire in 1 in 25 persons of European descent (1 in 2000 persons of European descent land, Beaumont Hospital, Dublin (N.G.M.); and UCL Respiratory, Division of Medi are homozygotes). Mild AAT deficiency typically results from a different amino cine, Rayne Institute, University College acid replacement, Glu264Val (the S allele), which is found in 1 in 4 persons in the London, London (D.A.L.). Address re Iberian peninsula. However, many other alleles have been described that have vari- print requests to Dr. Lomas at UCL Re spiratory, Rayne Institute, University Col able effects, such as a lack of protein production (null alleles), production of mis- lege London, London WC1E 6JF, United folded protein, or no effect on the level or function of circulating AAT (Table 1). Kingdom, or at d . -

Efficacy and Safety of Human Serum Albumin–Cisplatin Complex In
International Journal of Molecular Sciences Article Efficacy and Safety of Human Serum Albumin–Cisplatin Complex in U87MG Xenograft Mouse Models Cho Rong Park 1,2,3, Hyo Young Kim 1,2, Myung Geun Song 1,2,4,*, Yun-Sang Lee 1,5, Hyewon Youn 1,2,6, June-Key Chung 1,2,3,4, Gi Jeong Cheon 1,2,4,5,7 and Keon Wook Kang 1,2,3,4,5,6,8,* 1 Department of Nuclear Medicine, Seoul National University Hospital, 101 Daehak-ro, Jongno-gu, Seoul 03080, Korea; siofl[email protected] (C.R.P.); [email protected] (H.Y.K.); [email protected] (Y.-S.L.); [email protected] (H.Y.); [email protected] (J.-K.C.); [email protected] (G.J.C.) 2 Laboratory of Molecular Imaging and Therapy, Cancer Research Institute, Seoul 03080, Korea 3 Department of Biomedical Sciences, Seoul National University Graduate School, Biomedical Science Building, Seoul 03080, Korea 4 Biomedical Research Institute, Seoul National University Hospital, Seoul 03080, Korea 5 Radiation Research Institute, Seoul National University College of Medicine, Seoul 03080, Korea 6 Cancer Imaging Center, Seoul National University Hospital, Seoul 03080, Korea 7 Institute on Aging, Seoul National University, Seoul 08826, Korea 8 Bio-MAX Institute, Seoul National University, Seoul 08826, Korea * Correspondence: [email protected] (M.G.S.); [email protected] (K.W.K.); Tel.: +82-2-2072-2803 (K.W.K.) Received: 18 September 2020; Accepted: 22 October 2020; Published: 26 October 2020 Abstract: Cisplatin (cis-diamminedichloroplatinum (II), CDDP) is a chemotherapeutic drug widely used against many solid tumors. -

Identification of Key Pathways and Genes in Dementia Via Integrated Bioinformatics Analysis
bioRxiv preprint doi: https://doi.org/10.1101/2021.04.18.440371; this version posted July 19, 2021. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. Identification of Key Pathways and Genes in Dementia via Integrated Bioinformatics Analysis Basavaraj Vastrad1, Chanabasayya Vastrad*2 1. Department of Biochemistry, Basaveshwar College of Pharmacy, Gadag, Karnataka 582103, India. 2. Biostatistics and Bioinformatics, Chanabasava Nilaya, Bharthinagar, Dharwad 580001, Karnataka, India. * Chanabasayya Vastrad [email protected] Ph: +919480073398 Chanabasava Nilaya, Bharthinagar, Dharwad 580001 , Karanataka, India bioRxiv preprint doi: https://doi.org/10.1101/2021.04.18.440371; this version posted July 19, 2021. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. Abstract To provide a better understanding of dementia at the molecular level, this study aimed to identify the genes and key pathways associated with dementia by using integrated bioinformatics analysis. Based on the expression profiling by high throughput sequencing dataset GSE153960 derived from the Gene Expression Omnibus (GEO), the differentially expressed genes (DEGs) between patients with dementia and healthy controls were identified. With DEGs, we performed a series of functional enrichment analyses. Then, a protein–protein interaction (PPI) network, modules, miRNA-hub gene regulatory network and TF-hub gene regulatory network was constructed, analyzed and visualized, with which the hub genes miRNAs and TFs nodes were screened out. Finally, validation of hub genes was performed by using receiver operating characteristic curve (ROC) analysis. -
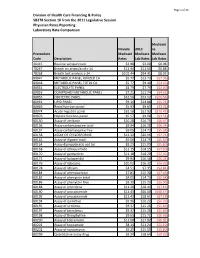
2019 Laboratory Report
Page 1 of 24 Division of Health Care Financing & Policy SB278 Section 16 from the 2011 Legislative Session Physician Rates Reporting Laboratory Rate Comparison Medicaid Nevada 2019 vs. Proccedure Medicaid Medicare Medicare Code Description Rates Lab Rates Lab Rates 36415 Routine venipuncture $3.98 $3.00 $0.98 78267 Breath tst attain/anal c-14 $11.94 $11.06 $0.88 78268 Breath test analysis c-14 $102.44 $94.41 $8.03 80047 METABOLIC PANEL IONIZED CA $5.77 $13.73 ($7.96) 80048 METABOLIC PANEL TOTAL CA $5.77 $9.40 ($3.63) 80051 ELECTROLYTE PANEL $4.79 $7.79 ($3.00) 80053 COMPREHEN METABOLIC PANEL $7.21 $11.74 ($4.53) 80055 OBSTETRIC PANEL $32.56 $53.12 ($20.56) 80061 LIPID PANEL $9.14 $14.88 ($5.74) 80069 Renal function panel $5.93 $9.65 ($3.72) 80074 Acute hepatitis panel $32.50 $52.93 ($20.43) 80076 Hepatic function panel $5.57 $9.08 ($3.51) 80150 Assay of amikacin $10.28 $16.75 ($6.47) 80156 Assay carbamazepine total $9.94 $16.18 ($6.24) 80157 Assay carbamazepine free $9.05 $14.73 ($5.68) 80158 ASSAY OF CYCLOSPORINE $12.32 $20.06 ($7.74) 80162 Assay of digoxin total $9.06 $14.75 ($5.69) 80164 Assay dipropylacetic acd tot $9.25 $15.05 ($5.80) 80168 Assay of ethosuximide $11.15 $18.15 ($7.00) 80170 Assay of gentamicin $11.18 $18.20 ($7.02) 80173 Assay of haloperidol $9.94 $16.18 ($6.24) 80176 Assay of lidocaine $10.02 $16.32 ($6.30) 80178 Assay of lithium $4.51 $7.35 ($2.84) 80184 Assay of phenobarbital $7.81 $15.30 ($7.49) 80185 Assay of phenytoin total $9.05 $14.73 ($5.68) 80186 Assay of phenytoin free $9.39 $15.29 ($5.90) 80188 Assay of -

The Influence of Oxidative Stress on Serum Albumin Structure As A
pharmaceuticals Article The Influence of Oxidative Stress on Serum Albumin Structure as a Carrier of Selected Diazaphenothiazine with Potential Anticancer Activity Małgorzata Maci ˛azek-Jurczyk˙ 1,* , Beata Morak-Młodawska 2 , Małgorzata Jele ´n 2 , Wiktoria Kope´c 1, Agnieszka Szkudlarek 1 , Aleksandra Owczarzy 1, Karolina Kulig 1, Wojciech Rogóz˙ 1 and Jadwiga Pozycka˙ 1 1 Department of Physical Pharmacy, Faculty of Pharmaceutical Sciences in Sosnowiec, Medical University of Silesia in Katowice, 40-055 Katowice, Poland; [email protected] (W.K.); [email protected] (A.S.); [email protected] (A.O.); [email protected] (K.K.); [email protected] (W.R.); [email protected] (J.P.) 2 Department of Organic Chemistry, Faculty of Pharmaceutical Sciences in Sosnowiec, Medical University of Silesia in Katowice, 40-055 Katowice, Poland; [email protected] (B.M.-M.); [email protected] (M.J.) * Correspondence: [email protected]; Tel.: +48-32-364-15-80 Abstract: Albumin is one of the most important proteins in human blood. Among its multiple functions, drug binding is crucial in terms of drug distribution in human body. This protein undergoes many modifications that are certain to influence protein activity and affect its structure. One such reaction is albumin oxidation. Chloramine T is a strong oxidant. Solutions of human serum albumin, both non-modified and modified by chloramine T, were examined with the use of fluorescence, ˙ Citation: Maci ˛azek-Jurczyk, M.; absorption and circular dichroism (CD) spectroscopy. 10H-3,6-diazaphenothiazine (DAPT) has Morak-Młodawska, B.; Jele´n,M.; anticancer activity and it has been studied for the first time in terms of binding with human serum Kope´c,W.; Szkudlarek, A.; Owczarzy, albumin—its potential as a transporting protein. -

Detection of Food Proteins in Human Serum Using Mass Spectrometry Methods
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Dissertations, Theses, & Student Research in Food Science and Technology Food Science and Technology Department 8-2020 DETECTION OF FOOD PROTEINS IN HUMAN SERUM USING MASS SPECTROMETRY METHODS Abigail S. Burrows University of Nebraska-Lincoln, [email protected] Follow this and additional works at: https://digitalcommons.unl.edu/foodscidiss Part of the Food Science Commons Burrows, Abigail S., "DETECTION OF FOOD PROTEINS IN HUMAN SERUM USING MASS SPECTROMETRY METHODS" (2020). Dissertations, Theses, & Student Research in Food Science and Technology. 109. https://digitalcommons.unl.edu/foodscidiss/109 This Article is brought to you for free and open access by the Food Science and Technology Department at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Dissertations, Theses, & Student Research in Food Science and Technology by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. DETECTION OF FOOD PROTEINS IN HUMAN SERUM USING MASS SPECTROMETRY METHODS by Abigail S. Burrows A DISSERTATION Presented to the Faculty of The Graduate College of the University of Nebraska In Partial Fulfillment of Requirements For the Degree of Doctor of Philosophy Major: Food Science and Technology Under the Supervision of Professor Philip E. Johnson Lincoln, Nebraska August, 2020 DETECTION OF FOOD PROTEINS IN HUMAN SERUM USING MASS SPECTROMETRY METHODS Abigail S. Burrows, Ph.D. University of Nebraska, 2020 Advisor: Philip E. Johnson Allergenic peanut proteins are highly resistant to digestion and are detectable by immunoassays after gastrointestinal digestion. The application of liquid chromatography-tandem mass spectrometry (LC-MS/MS) methods for in vivo detection of peptides originating from allergenic food proteins has not been thoroughly studied.