Meningiomas and Schwannomas in Neurofibromatosis 2
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Neurofibromatosis Type 2 (NF2)
International Journal of Molecular Sciences Review Neurofibromatosis Type 2 (NF2) and the Implications for Vestibular Schwannoma and Meningioma Pathogenesis Suha Bachir 1,† , Sanjit Shah 2,† , Scott Shapiro 3,†, Abigail Koehler 4, Abdelkader Mahammedi 5 , Ravi N. Samy 3, Mario Zuccarello 2, Elizabeth Schorry 1 and Soma Sengupta 4,* 1 Department of Genetics, Cincinnati Children’s Hospital, Cincinnati, OH 45229, USA; [email protected] (S.B.); [email protected] (E.S.) 2 Department of Neurosurgery, University of Cincinnati, Cincinnati, OH 45267, USA; [email protected] (S.S.); [email protected] (M.Z.) 3 Department of Otolaryngology, University of Cincinnati, Cincinnati, OH 45267, USA; [email protected] (S.S.); [email protected] (R.N.S.) 4 Department of Neurology, University of Cincinnati, Cincinnati, OH 45267, USA; [email protected] 5 Department of Radiology, University of Cincinnati, Cincinnati, OH 45267, USA; [email protected] * Correspondence: [email protected] † These authors contributed equally. Abstract: Patients diagnosed with neurofibromatosis type 2 (NF2) are extremely likely to develop meningiomas, in addition to vestibular schwannomas. Meningiomas are a common primary brain tumor; many NF2 patients suffer from multiple meningiomas. In NF2, patients have mutations in the NF2 gene, specifically with loss of function in a tumor-suppressor protein that has a number of synonymous names, including: Merlin, Neurofibromin 2, and schwannomin. Merlin is a 70 kDa protein that has 10 different isoforms. The Hippo Tumor Suppressor pathway is regulated upstream by Merlin. This pathway is critical in regulating cell proliferation and apoptosis, characteristics that are important for tumor progression. -

Risk Factors for Gliomas and Meningiomas in Males in Los Angeles County1
[CANCER RESEARCH 49, 6137-6143. November 1, 1989] Risk Factors for Gliomas and Meningiomas in Males in Los Angeles County1 Susan Preston-Martin,2 Wendy Mack, and Brian E. Henderson Department of Preventive Medicine, University of Southern California School of Medicine, Los Angeles, California 90033 ABSTRACT views with proxy respondents, we were unable to include a large proportion of otherwise eligible cases because they were deceased or Detailed job histories and information about other suspected risk were too ill or impaired to participate in an interview. The Los Angeles factors were obtained during interviews with 272 men aged 25-69 with a County Cancer Surveillance Program identified the cases (26). All primary brain tumor first diagnosed during 1980-1984 and with 272 diagnoses had been microscopically confirmed. individually matched neighbor controls. Separate analyses were con A total of 478 patients were identified. The hospital and attending ducted for the 202 glioma pairs and the 70 meningioma pairs. Meningi- physician granted us permission to contact 396 (83%) patients. We oma, but not glioma, was related to having a serious head injury 20 or were unable to locate 22 patients, 38 chose not to participate, and 60 more years before diagnosis (odds ratio (OR) = 2.3; 95% confidence were aphasie or too ill to complete the interview. We interviewed 277 interval (CI) = 1.1-5.4), and a clear dose-response effect was observed patients (74% of the 374 patients contacted about the study or 58% of relating meningioma risk to number of serious head injuries (/' for trend the initial 478 patients). -

Information About Mosaic Neurofibromatosis Type 2 (NF2)
Information about mosaic Neurofibromatosis type 2 (NF2) NF2 occurs because of a mutation (change) in the NF2 gene. When this change is present at the time of conception the changed gene will be present in all the cells of the baby. When this mutation occurs later in the development of the forming embryo, the baby will go on to have a mix of cells: some with the “normal” genetic information and some with the changed information. This mix of cells is called mosaicism. Approximately half the people who have a diagnosis of NF2 have inherited the misprinted NF2 gene change from their mother or father who will also have NF2. They will have that misprinted gene in all the cells of their body. When they have their children, there will be a 1 in 2 chance of passing on NF2 to each child they have. However about half of people with NF2 are the first person in the family to be affected. They have no family history and have not inherited the condition from a parent. When doctors studied this group of patients more closely they noticed certain characteristics. Significantly they observed that fewer children had inherited NF2 than expected some people in this group had relatively mild NF2 NF2 tumours in some patients tended to grow on one side of their body rather than both sides that when a blood sample was tested to identify the NF2 gene, the gene change could not be found in 30-40% of people This lead researchers to conclude that this group of people were most likely to be mosaic for NF2 i.e. -

Brain Tumors
BRAIN TUMORS What kinds of brain tumors affect pets? Brain tumors occur relatively often in dogs and cats. The most common type of brain tumor is a meningioma, which originates from the layer surrounding the brain, called the meninges. Meningiomas are slow-growing benign tumors that are often present for months to years before clinical signs appear. Other common types in- clude glial cell tumors, which originate in the brain tissue, and choroid plexus tumors, which originate from the tissue in the brain that produces spinal fluid. Tumors in structures around the brain (like nasal tumors, skull tumors, and pituitary tumors) may also com- press the brain. Lymphoma is a type of cancer that can affect multiple parts of the brain and spine. What are the symptoms? Symptoms vary and depend on the size and location of the tumor. Common signs include changes in behavior, circling or pacing, staring into space, getting stuck in corners, and seizures. Other signs can include weakness, lack of alertness, difficulty eating or swallowing, and problems with the “vestibular system,” or system of bal- ance, such as lack of coordination, head tilt, leaning/circling/falling to one side, and abnormal eye movements. How are brain tumors diagnosed? An MRI scan produces an image of the brain that’s more detailed than a CT scan or x-ray and can identify a tumor. Often the appearance of the tumor on MRI suggests the type of tumor (i.e. meningioma vs lymphoma). However, a biopsy of the tumor is required to give a definitive diagnosis. In some cases, spinal fluid is collected to help diagnose lymphoma. -

Central Nervous System Tumors General ~1% of Tumors in Adults, but ~25% of Malignancies in Children (Only 2Nd to Leukemia)
Last updated: 3/4/2021 Prepared by Kurt Schaberg Central Nervous System Tumors General ~1% of tumors in adults, but ~25% of malignancies in children (only 2nd to leukemia). Significant increase in incidence in primary brain tumors in elderly. Metastases to the brain far outnumber primary CNS tumors→ multiple cerebral tumors. One can develop a very good DDX by just location, age, and imaging. Differential Diagnosis by clinical information: Location Pediatric/Young Adult Older Adult Cerebral/ Ganglioglioma, DNET, PXA, Glioblastoma Multiforme (GBM) Supratentorial Ependymoma, AT/RT Infiltrating Astrocytoma (grades II-III), CNS Embryonal Neoplasms Oligodendroglioma, Metastases, Lymphoma, Infection Cerebellar/ PA, Medulloblastoma, Ependymoma, Metastases, Hemangioblastoma, Infratentorial/ Choroid plexus papilloma, AT/RT Choroid plexus papilloma, Subependymoma Fourth ventricle Brainstem PA, DMG Astrocytoma, Glioblastoma, DMG, Metastases Spinal cord Ependymoma, PA, DMG, MPE, Drop Ependymoma, Astrocytoma, DMG, MPE (filum), (intramedullary) metastases Paraganglioma (filum), Spinal cord Meningioma, Schwannoma, Schwannoma, Meningioma, (extramedullary) Metastases, Melanocytoma/melanoma Melanocytoma/melanoma, MPNST Spinal cord Bone tumor, Meningioma, Abscess, Herniated disk, Lymphoma, Abscess, (extradural) Vascular malformation, Metastases, Extra-axial/Dural/ Leukemia/lymphoma, Ewing Sarcoma, Meningioma, SFT, Metastases, Lymphoma, Leptomeningeal Rhabdomyosarcoma, Disseminated medulloblastoma, DLGNT, Sellar/infundibular Pituitary adenoma, Pituitary adenoma, -

Adrenal Neuroblastoma Mimicking Pheochromocytoma in an Adult With
Khalayleh et al. Int Arch Endocrinol Clin Res 2017, 3:008 Volume 3 | Issue 1 International Archives of Endocrinology Clinical Research Case Report : Open Access Adrenal Neuroblastoma Mimicking Pheochromocytoma in an Adult with Neurofibromatosis Type 1 Harbi Khalayleh1, Hilla Knobler2, Vitaly Medvedovsky2, Edit Feldberg3, Judith Diment3, Lena Pinkas4, Guennadi Kouniavsky1 and Taiba Zornitzki2* 1Department of Surgery, Hebrew University Medical School of Jerusalem, Israel 2Endocrinology, Diabetes and Metabolism Institute, Kaplan Medical Center, Hebrew University Medical School of Jerusalem, Israel 3Pathology Institute, Kaplan Medical Center, Israel 4Nuclear Medicine Institute, Kaplan Medical Center, Israel *Corresponding author: Taiba Zornitzki, MD, Endocrinology, Diabetes and Metabolism Institute, Kaplan Medical Center, Hebrew University Medical School of Jerusalem, Bilu 1, 76100 Rehovot, Israel, Tel: +972-894- 41315, Fax: +972-8 944-1912, E-mail: [email protected] Context 2. This is the first reported case of an adrenal neuroblastoma occurring in an adult patient with NF1 presenting as a large Neurofibromatosis type 1 (NF1) is a genetic disorder asso- adrenal mass with increased catecholamine levels mimicking ciated with an increased risk of malignant disorders. Adrenal a pheochromocytoma. neuroblastoma is considered an extremely rare tumor in adults and was not previously described in association with NF1. 3. This case demonstrates the clinical overlap between pheo- Case description: A 42-year-old normotensive woman with chromocytoma and neuroblastoma. typical signs of NF1 underwent evaluation for abdominal pain, Keywords and a large 14 × 10 × 16 cm left adrenal mass displacing the Adrenal neuroblastoma, Neurofibromatosis type 1, Pheo- spleen, pancreas and colon was found. An initial diagnosis of chromocytoma, Neural crest-derived tumors pheochromocytoma was done based on the known strong association between pheochromocytoma, NF1 and increased catecholamine levels. -

Neuro-Oncology
Neuro-Oncology Neuro-Oncology 17:iv1–iv62, 2015. doi:10.1093/neuonc/nov189 CBTRUS Statistical Report: Primary Brain and Central Nervous System Tumors Diagnosed in the United States in 2008-2012 Quinn T. Ostrom M.A., M.P.H.1,2*, Haley Gittleman M.S.1,2*, Jordonna Fulop R.N.1, Max Liu3, Rachel Blanda4, Courtney Kromer B.A.5, Yingli Wolinsky Ph.D., M.B.A.1,2, Carol Kruchko B.A.2, and Jill S. Barnholtz-Sloan Ph.D.1,2 1Case Comprehensive Cancer Center, Case Western Reserve University School of Medicine, Cleveland, OH USA 2Central Brain Tumor Registry of the United States, Hinsdale, IL USA 3Solon High School, Solon, OH USA 4Georgetown University, Washington D.C. USA 5Case Western Reserve University School of Medicine, Cleveland, OH USA *Contributed equally to this Report. Introduction for collection of central (state) cancer data as mandated in 1992 by Public Law 102-515, the Cancer Registries Amendment The objective of the CBTRUS Statistical Report: Primary Brain and Act.2 This mandate was expanded to include non-malignant Central Nervous System Tumors Diagnosed in the United States brain tumors diagnosed in 2004 and later with the 2002 in 2008-2012 is to provide a comprehensive summary of the passage of Public Law 107–260.3 CBTRUS researchers combine current descriptive epidemiology of primary brain and central the NPCR data with data from the SEER program4 of the NCI, nervous system (CNS) tumors in the United States (US) popula- which was established for national cancer surveillance in the tion. CBTRUS obtained the latest available data on all newly early 1970s. -
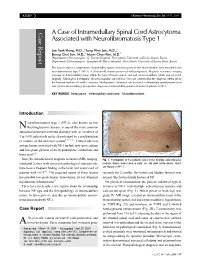
A Case of Intramedullary Spinal Cord Astrocytoma Associated with Neurofibromatosis Type 1
KISEP J Korean Neurosurg Soc 36 : 69-71, 2004 Case Report A Case of Intramedullary Spinal Cord Astrocytoma Associated with Neurofibromatosis Type 1 Jae Taek Hong, M.D.,1 Sang Won Lee, M.D.,1 Byung Chul Son, M.D.,1 Moon Chan Kim, M.D.2 Department of Neurosurgery,1 St. Vincent Hospital, The Catholic University of Korea, Suwon, Korea Department of Neurosurgery,2 Kangnam St. Mary's Hospital, The Catholic University of Korea, Seoul, Korea The authors report a symptomatic intramedullary spinal cord astrocytoma in the thoracolumbar area associated with neurofibromatosis type 1 (NF-1). A 38-year-old woman presented with paraparesis. Magnetic resonance imaging revealed an intramedullary lesion within the lower thoracic spinal cord and conus medullaris, which was removed surgically. Pathological investigation showed anaplastic astrocytoma. This case confirms that the diagnosis criteria set by the National Institute of Health Consensus Development Conference can be useful to differentiate ependymoma from astrocytoma when making a preoperative diagnosis of intramedullary spinal cord tumor in patients of NF-1. KEY WORDS : Astrocytoma·Intramedullary cord tumor·Neurofibromatosis. Introduction eurofibromatosis type 1 (NF-1), also known as von N Recklinghausen's disease, is one of the most common autosomal dominant inherited disorders with an incidence of 1 in 3,000 individuals and is characterized by a predisposition to tumors of the nervous system5,6,12,16). Central nervous system lesions associated with NF-1 include optic nerve glioma and low-grade gliomas of the hypothalamus, cerebellum and brain stem6,10). Since the introduction of magnetic resonance(MR) imaging, Fig. 1. Photograph of the patient's back shows multiple subcutaneous incidental lesions with uncertain pathological characteristic nodules (black arrow) and a cafe-au-lait spot (white arrow), which have been a frequent finding in the brain and spinal cord of are typical of NF-1. -

Surgical Results of the Resection of Spinal Meningioma with the Inner Layer of Dura More Than 10 Years After Surgery
www.nature.com/scientificreports OPEN Surgical results of the resection of spinal meningioma with the inner layer of dura more than 10 years after surgery Hiroyuki Tominaga1*, Ichiro Kawamura1, Kosei Ijiri2, Kazunori Yone1 & Noboru Taniguchi1 Most spinal meningiomas arise from the thoracic dura in middle-aged and elderly women. Simpson grade 1 resection is recommended to avoid recurrence. For ventral and ventrolateral tumors, reconstruction after total dural resection is difcult, and spinal fuid leakage is likely. To overcome this concern, Saito et al. developed the technique of resecting the tumor with the inner dural layer, preserving the outer dural layer. Although meningioma rarely recurs, the recurrence period is approximately 8 years postoperatively. No studies have evaluated long-term (> 10-year) outcomes of the Saito method. Here, we report 10 cases of the Saito method with > 10-year follow-up and compare outcomes with those of other standard approaches. Twenty-nine pathology-confrmed meningioma patients underwent surgery in our department, ten with the Saito method. We investigated resection method (dura mater treatment), pathological type, and recurrence and compared pre- and postoperative clinical fndings. The median follow-up was 132 months. Recurrence occurred after Simpson grades 3 and 4 resection. Simpson grades 1, 2, and the Saito method resulted in no recurrence. Neurological symptoms improved in all patients at fnal follow-up. This is the frst report of long-term outcomes of the Saito method. The method achieved good neurological improvement with no recurrence in > 10-year follow-up. Spinal cord meningioma is an intradural, extramedullary tumor that occurs most frequently at the thoracic level in middle-aged and elderly women. -

Inhibition of Mir-1193 Leads to Synthetic Lethality in Glioblastoma
Zhang et al. Cell Death and Disease (2020) 11:602 https://doi.org/10.1038/s41419-020-02812-3 Cell Death & Disease ARTICLE Open Access Inhibition of miR-1193 leads to synthetic lethality in glioblastoma multiforme cells deficient of DNA-PKcs Jing Zhang1,LiJing1,SubeeTan2, Er-Ming Zeng3, Yingbo Lin4, Lingfeng He 1, Zhigang Hu 1, Jianping Liu1 and Zhigang Guo1 Abstract Glioblastoma multiforme (GBM) is the most malignant primary brain tumor and has the highest mortality rate among cancers and high resistance to radiation and cytotoxic chemotherapy. Although some targeted therapies can partially inhibit oncogenic mutation-driven proliferation of GBM cells, therapies harnessing synthetic lethality are ‘coincidental’ treatments with high effectiveness in cancers with gene mutations, such as GBM, which frequently exhibits DNA-PKcs mutation. By implementing a highly efficient high-throughput screening (HTS) platform using an in-house-constructed genome-wide human microRNA inhibitor library, we demonstrated that miR-1193 inhibition sensitized GBM tumor cells with DNA-PKcs deficiency. Furthermore, we found that miR-1193 directly targets YY1AP1, leading to subsequent inhibition of FEN1, an important factor in DNA damage repair. Inhibition of miR-1193 resulted in accumulation of DNA double-strand breaks and thus increased genomic instability. RPA-coated ssDNA structures enhanced ATR checkpoint kinase activity, subsequently activating the CHK1/p53/apoptosis axis. These data provide a preclinical theory for the application of miR-1193 inhibition as a potential synthetic lethal approach targeting GBM cancer cells with DNA-PKcs fi 1234567890():,; 1234567890():,; 1234567890():,; 1234567890():,; de ciency. Introduction In response to DNA damage, cells activate the DNA Glioblastoma multiforme (GBM), exhibits highly damage response (DDR) network, allowing DNA repair aggressive invasion, a high mortality rate, and high resis- through the regulation of cell-cycle progression, DNA tance to radiation and cytotoxic chemotherapy, and thus damage repair or apoptosis4. -

The WHO Grade I Collagen-Forming Meningioma Produces Angiogenic Substances
ANTICANCER RESEARCH 36: 941-950 (2016) The WHO Grade I Collagen-forming Meningioma Produces Angiogenic Substances. A New Meningioma Entity JOHANNES HAYBAECK1*, ELISABETH SMOLLE1,2*, BERNADETTE SCHÖKLER3 and REINHOLD KLEINERT1 1Department of Neuropathology, Institute of Pathology, Medical University Graz, Graz, Austria; 2Department of Internal Medicine, Division of Pulmonology, Medical University Graz, Graz, Austria; 3Department of Neurosurgery, Medical University Graz, Graz, Austria Abstract. Background: Meningiomas arise from arachnoid seems to be most appropriate. Conclusion: The "WHO Grade cap cells, the so-called meningiothelial cells. They account I collagen-forming meningioma" reported herein produces for 20-36% of all primary intracranial tumours, and arise collagen and angiogenic substances. To the best of our with an annual incidence of 1.8-13 per 100,000 knowledge, no such entity has been reported on in previous individuals/year. According to their histopathological features literature. We propose this collagen-producing meningioma meningiomas are classified either as grade I (meningiothelial, as a novel WHO grade I meningioma sub-type. fibrous/fibroblastic, transitional/mixed, psammomatous, angiomatous, microcystic, secretory and the Meningiomas arise from arachnoid cap cells, the so-called lympholasmacyterich sub-type), grade II (atypical and clear- meningiothelial cells (1-5). They account for 20-36% of all cell sub-type) or grade III (malignant or anaplastic primary intracranial tumours, and arise with an annual phenotype). Case Report: A 62-year-old female patient incidence of 1.8-13 per 100,000 individuals/year (1-5). presented to the hospital because of progressive obliviousness Meningiomas are the second most common central nervous and concentration difficulties. In the magnetic resonance system neoplasms in adults (6-9). -

A Case of Mushroom‑Shaped Anaplastic Oligodendroglioma Resembling Meningioma and Arteriovenous Malformation: Inadequacies of Diagnostic Imaging
EXPERIMENTAL AND THERAPEUTIC MEDICINE 10: 1499-1502, 2015 A case of mushroom‑shaped anaplastic oligodendroglioma resembling meningioma and arteriovenous malformation: Inadequacies of diagnostic imaging YAOLING LIU1,2, KANG YANG1, XU SUN1, XINYU LI1, MINGHAI WEI1, XIANG'EN SHI2, NINGWEI CHE1 and JIAN YIN1 1Department of Neurosurgery, The Second Affiliated Hospital of Dalian Medical University, Dalian, Liaoning 116044; 2Department of Neurosurgery, Affiliated Fuxing Hospital, The Capital University of Medical Sciences, Beijing 100038, P.R. China Received December 29, 2014; Accepted June 29, 2015 DOI: 10.3892/etm.2015.2676 Abstract. Magnetic resonance imaging (MRI) is the most tomas (WHO IV) (2). The median survival times of patients widely discussed and clinically employed method for the with WHO II and WHO III oligodendrogliomas are 9.8 and differential diagnosis of oligodendrogliomas; however, 3.9 years, respectively (1,2), and 6.3 and 2.8 years, respec- MRI occasionally produces unclear results that can hinder tively, if mixed with astrocytes (3,4). Surgical excision and a definitive oligodendroglioma diagnosis. The present study postoperative adjuvant radiotherapy is the traditional therapy describes the case of a 34-year-old man that suffered from for oligodendroglioma; however, studies have observed that, headache and right upper‑extremity weakness for 2 months. among intracranial tumors, anaplastic oligodendrogliomas Based on the presurgical evaluation, it was suggested that the are particularly sensitive to chemotherapy, and the prognosis patient had a World Health Organization (WHO) grade II-II of patients treated with chemotherapy is more favorable glioma, meningioma or arteriovenous malformation (AVM), than that of patients treated with radiotherapy (5‑7).