Breeding Behaviour of the Brown Goshawk Accipiter Fasciatus by T
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Australian Diurnal Raptors and Airports
Australian diurnal raptors and airports Photo: John Barkla, BirdLife Australia William Steele Australasian Raptor Association BirdLife Australia Australian Aviation Wildlife Hazard Group Forum Brisbane, 25 July 2013 So what is a raptor? Small to very large birds of prey. Diurnal, predatory or scavenging birds. Sharp, hooked bills and large powerful feet with talons. Order Falconiformes: 27 species on Australian list. Family Falconidae – falcons/ kestrels Family Accipitridae – eagles, hawks, kites, osprey Falcons and kestrels Brown Falcon Black Falcon Grey Falcon Nankeen Kestrel Australian Hobby Peregrine Falcon Falcons and Kestrels – conservation status Common Name EPBC Qld WA SA FFG Vic NSW Tas NT Nankeen Kestrel Brown Falcon Australian Hobby Grey Falcon NT RA Listed CR VUL VUL Black Falcon EN Peregrine Falcon RA Hawks and eagles ‐ Osprey Osprey Hawks and eagles – Endemic hawks Red Goshawk female Hawks and eagles – Sparrowhawks/ goshawks Brown Goshawk Photo: Rik Brown Hawks and eagles – Elanus kites Black‐shouldered Kite Letter‐winged Kite ~ 300 g Hover hunters Rodent specialists LWK can be crepuscular Hawks and eagles ‐ eagles Photo: Herald Sun. Hawks and eagles ‐ eagles Large ‐ • Wedge‐tailed Eagle (~ 4 kg) • Little Eagle (< 1 kg) • White‐bellied Sea‐Eagle (< 4 kg) • Gurney’s Eagle Scavengers of carrion, in addition to hunters Fortunately, mostly solitary although some multiple strikes on aircraft Hawks and eagles –large kites Black Kite Whistling Kite Brahminy Kite Frequently scavenge Large at ~ 600 to 800 g BK and WK flock and so high risk to aircraft Photo: Jill Holdsworth Identification Beruldsen, G (1995) Raptor Identification. Privately published by author, Kenmore Hills, Queensland, pp. 18‐19, 26‐27, 36‐37. -

Common Birds in Tilligerry Habitat
Common Birds in Tilligerry Habitat Dedicated bird enthusiasts have kindly contributed to this sequence of 106 bird species spotted in the habitat over the last few years Kookaburra Red-browed Finch Black-faced Cuckoo- shrike Magpie-lark Tawny Frogmouth Noisy Miner Spotted Dove [1] Crested Pigeon Australian Raven Olive-backed Oriole Whistling Kite Grey Butcherbird Pied Butcherbird Australian Magpie Noisy Friarbird Galah Long-billed Corella Eastern Rosella Yellow-tailed black Rainbow Lorikeet Scaly-breasted Lorikeet Cockatoo Tawny Frogmouth c Noeline Karlson [1] ( ) Common Birds in Tilligerry Habitat Variegated Fairy- Yellow Faced Superb Fairy-wren White Cheeked Scarlet Honeyeater Blue-faced Honeyeater wren Honeyeater Honeyeater White-throated Brown Gerygone Brown Thornbill Yellow Thornbill Eastern Yellow Robin Silvereye Gerygone White-browed Eastern Spinebill [2] Spotted Pardalote Grey Fantail Little Wattlebird Red Wattlebird Scrubwren Willie Wagtail Eastern Whipbird Welcome Swallow Leaden Flycatcher Golden Whistler Rufous Whistler Eastern Spinebill c Noeline Karlson [2] ( ) Common Sea and shore birds Silver Gull White-necked Heron Little Black Australian White Ibis Masked Lapwing Crested Tern Cormorant Little Pied Cormorant White-bellied Sea-Eagle [3] Pelican White-faced Heron Uncommon Sea and shore birds Caspian Tern Pied Cormorant White-necked Heron Great Egret Little Egret Great Cormorant Striated Heron Intermediate Egret [3] White-bellied Sea-Eagle (c) Noeline Karlson Uncommon Birds in Tilligerry Habitat Grey Goshawk Australian Hobby -

The Conservation Ecology of the Endangered Numbat Myrmecobius Fasciatus (Marsupialia: Myrmecobiidae) Reintroduced to Scotia and Yookamurra Sanctuaries, Australia
Numbat nirvana: the conservation ecology of the endangered numbat ANGOR UNIVERSITY Myrmecobius fasciatus (Marsupialia: Myrmecobiidae) reintroduced to Scotia and Yookamurra Sanctuaries, Australia Hayward, M.W.; Poh, A.S.; Cathcart, J.; Churcher, C.; Bentley, J.; Herman, K.; Kemp, L.; Riessen, N.; Scully, P.; Dion, C.H.; Legge, S.; Carter, A.; Gibb, H.; Friend, J.A. Australian Journal of Zoology DOI: PRIFYSGOL BANGOR / B 10.1071/ZO15028 Published: 15/10/2015 Peer reviewed version Cyswllt i'r cyhoeddiad / Link to publication Dyfyniad o'r fersiwn a gyhoeddwyd / Citation for published version (APA): Hayward, M. W., Poh, A. S., Cathcart, J., Churcher, C., Bentley, J., Herman, K., Kemp, L., Riessen, N., Scully, P., Dion, C. H., Legge, S., Carter, A., Gibb, H., & Friend, J. A. (2015). Numbat nirvana: the conservation ecology of the endangered numbat Myrmecobius fasciatus (Marsupialia: Myrmecobiidae) reintroduced to Scotia and Yookamurra Sanctuaries, Australia. Australian Journal of Zoology. https://doi.org/10.1071/ZO15028 Hawliau Cyffredinol / General rights Copyright and moral rights for the publications made accessible in the public portal are retained by the authors and/or other copyright owners and it is a condition of accessing publications that users recognise and abide by the legal requirements associated with these rights. • Users may download and print one copy of any publication from the public portal for the purpose of private study or research. • You may not further distribute the material or use it for any profit-making activity or commercial gain • You may freely distribute the URL identifying the publication in the public portal ? Take down policy If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim. -

The Birds (Aves) of Oromia, Ethiopia – an Annotated Checklist
European Journal of Taxonomy 306: 1–69 ISSN 2118-9773 https://doi.org/10.5852/ejt.2017.306 www.europeanjournaloftaxonomy.eu 2017 · Gedeon K. et al. This work is licensed under a Creative Commons Attribution 3.0 License. Monograph urn:lsid:zoobank.org:pub:A32EAE51-9051-458A-81DD-8EA921901CDC The birds (Aves) of Oromia, Ethiopia – an annotated checklist Kai GEDEON 1,*, Chemere ZEWDIE 2 & Till TÖPFER 3 1 Saxon Ornithologists’ Society, P.O. Box 1129, 09331 Hohenstein-Ernstthal, Germany. 2 Oromia Forest and Wildlife Enterprise, P.O. Box 1075, Debre Zeit, Ethiopia. 3 Zoological Research Museum Alexander Koenig, Centre for Taxonomy and Evolutionary Research, Adenauerallee 160, 53113 Bonn, Germany. * Corresponding author: [email protected] 2 Email: [email protected] 3 Email: [email protected] 1 urn:lsid:zoobank.org:author:F46B3F50-41E2-4629-9951-778F69A5BBA2 2 urn:lsid:zoobank.org:author:F59FEDB3-627A-4D52-A6CB-4F26846C0FC5 3 urn:lsid:zoobank.org:author:A87BE9B4-8FC6-4E11-8DB4-BDBB3CFBBEAA Abstract. Oromia is the largest National Regional State of Ethiopia. Here we present the first comprehensive checklist of its birds. A total of 804 bird species has been recorded, 601 of them confirmed (443) or assumed (158) to be breeding birds. At least 561 are all-year residents (and 31 more potentially so), at least 73 are Afrotropical migrants and visitors (and 44 more potentially so), and 184 are Palaearctic migrants and visitors (and eight more potentially so). Three species are endemic to Oromia, 18 to Ethiopia and 43 to the Horn of Africa. 170 Oromia bird species are biome restricted: 57 to the Afrotropical Highlands biome, 95 to the Somali-Masai biome, and 18 to the Sudan-Guinea Savanna biome. -

South Africa: Magoebaskloof and Kruger National Park Custom Tour Trip Report
SOUTH AFRICA: MAGOEBASKLOOF AND KRUGER NATIONAL PARK CUSTOM TOUR TRIP REPORT 24 February – 2 March 2019 By Jason Boyce This Verreaux’s Eagle-Owl showed nicely one late afternoon, puffing up his throat and neck when calling www.birdingecotours.com [email protected] 2 | TRIP REPORT South Africa: Magoebaskloof and Kruger National Park February 2019 Overview It’s common knowledge that South Africa has very much to offer as a birding destination, and the memory of this trip echoes those sentiments. With an itinerary set in one of South Africa’s premier birding provinces, the Limpopo Province, we were getting ready for a birding extravaganza. The forests of Magoebaskloof would be our first stop, spending a day and a half in the area and targeting forest special after forest special as well as tricky range-restricted species such as Short-clawed Lark and Gurney’s Sugarbird. Afterwards we would descend the eastern escarpment and head into Kruger National Park, where we would make our way to the northern sections. These included Punda Maria, Pafuri, and the Makuleke Concession – a mouthwatering birding itinerary that was sure to deliver. A pair of Woodland Kingfishers in the fever tree forest along the Limpopo River Detailed Report Day 1, 24th February 2019 – Transfer to Magoebaskloof We set out from Johannesburg after breakfast on a clear Sunday morning. The drive to Polokwane took us just over three hours. A number of birds along the way started our trip list; these included Hadada Ibis, Yellow-billed Kite, Southern Black Flycatcher, Village Weaver, and a few brilliant European Bee-eaters. -

Australian Magpies Vary Mobbing Strategies, Not Intensity, Relative to Different Species of Predator
Clever strategists: Australian Magpies vary mobbing strategies, not intensity, relative to diVerent species of predator A KoboroV, G Kaplan and LJ Rogers Centre for Neuroscience and Animal Behaviour, University of New England, Australia ABSTRACT Anti-predator behaviour of magpies was investigated, using five species of model predators, at times of raising oVspring. We predicted diVerences in mobbing strate- gies for each predator presented and also that raising juveniles would aVect intensity of the mobbing event. Fourteen permanent resident family groups were tested using 5 diVerent types of predator (avian and reptilian) known to be of varying degrees of risk to magpies and common in their habitat. In all, 210 trials were conducted (across three diVerent stages of juvenile development). We found that the stage of juvenile development did not alter mobbing behaviour significantly, but predator type did. Aerial strategies (such as swooping) were elicited by taxidermic models of raptors, whereas a taxidermic model of a monitor lizard was approached on the ground and a model snake was rarely approached. Swooping patterns also changed according to which of the three raptors was presented. Our results show that, in contrast to findings in other species, magpies vary mobbing strategy depending on the predator rather than varying mobbing intensity. Subjects Animal Behavior, Zoology Keywords Risk assessment, Mobbing, Anti-predator behaviour, Australian Magpie INTRODUCTION Submitted 3 December 2012 For eVective defence against predation, potential prey must be able to accurately and Accepted 3 March 2013 Published 19 March 2013 rapidly assess risks and react appropriately. The level of intensity of any anti-predator responses may be associated with assessment of a number of factors. -

Crow Threat to Raptors?
PIED pirates CROW THREAT TO RAPTORS? There has been much debate about the effects on biodiversity of increasing Pied Crow and other corvid populations in Africa, but little has been quantified. It is evident from the latest bird atlas data that there are now more Pied Crows in southern Africa than there were a decade ago and that the species has spread into areas of the Karoo where it did not occur previously. But what effect is this having on other bird species? TEXT BY ROB SIMMONS & PHOEBE BARNARD ALBERT FRONEMAN arious reports indicate that one a Rock Kestrel and another a Southern crows are impacting negatively Pale Chanting Goshawk, all of which were on other creatures in their en- carrying mice in their talons. Prey-carrying vironment. Farmers complain raptors are usually provisioning dependent aboutV increasing predation by ‘crows’ on females or nestlings, so piracy affects more lambing ewes, birders suggest that more at- than simply the bird with the food. tacks are occurring on passerine birds, and The following three incidents serve to conservationists and members of the pub- highlight the modus operandi of the crows. lic report greater numbers of crows killing We then investigated other parts of Africa small tortoises. However, almost nothing to determine if piracy targeting raptors is has been quantified as to the level of such limited to western South Africa as a zone predation, the magnitude of the upsurge of Pied Crow range expansion or is a more in crow interactions and the effect on the widespread strategy. species being attacked. We do not yet have In the West Coast National Park in such data, but we do take a first look at 2008, our attention was drawn to a Black- what may be an increasing and somewhat shouldered Kite that was calling from a surprising addition to the negative effects height of over 150 metres and circling up- of crows – those on birds of prey. -

Exploring the Maintenance of Plumage Polymorphism in the Black Sparrowhawk
EXPLORING THE MAINTENANCE OF PLUMAGE POLYMORPHISM IN THE BLACK SPARROWHAWK GARETH JOHN TATE UniversityTTXGAR001 of Cape Town July 2016 SUBMITTED TO THE UNIVERSITY OF CAPE TOWN In fulfilment of the requirements for the degree of Doctor of Philosophy Percy FitzPatrick Institute of African Ornithology Department of Biological Sciences, Faculty of Science University of Cape Town SUPERVISED BY: DR. ARJUN AMAR & DR. JACQUELINE BISHOP The copyright of this thesis vests in the author. No quotation from it or information derived from it is to be published without full acknowledgement of the source. The thesis is to be used for private study or non- commercial research purposes only. Published by the University of Cape Town (UCT) in terms of the non-exclusive license granted to UCT by the author. University of Cape Town CONTENTS ABSTRACT------------------------------------------------------------------------------------------------------------------------------------------------------------------------------- 1 ACKNOWLEDGEMENTS------------------------------------------------------------------------------------------------------------------------------------------------- 4 CHAPTER 1: Introduction--------------------------------------------------------------------------------------------------------------------------------------------------------------------------------- 8 CHAPTER 2: Trait divergence in the presence of gene flow? A mitochondrial perspective on trait variation and colonisation of the Cape Peninsula by the black sparrowhawk Accipiter -

South Africa Mega Birding III 5Th to 27Th October 2019 (23 Days) Trip Report
South Africa Mega Birding III 5th to 27th October 2019 (23 days) Trip Report The near-endemic Gorgeous Bushshrike by Daniel Keith Danckwerts Tour leader: Daniel Keith Danckwerts Trip Report – RBT South Africa – Mega Birding III 2019 2 Tour Summary South Africa supports the highest number of endemic species of any African country and is therefore of obvious appeal to birders. This South Africa mega tour covered virtually the entire country in little over a month – amounting to an estimated 10 000km – and targeted every single endemic and near-endemic species! We were successful in finding virtually all of the targets and some of our highlights included a pair of mythical Hottentot Buttonquails, the critically endangered Rudd’s Lark, both Cape, and Drakensburg Rockjumpers, Orange-breasted Sunbird, Pink-throated Twinspot, Southern Tchagra, the scarce Knysna Woodpecker, both Northern and Southern Black Korhaans, and Bush Blackcap. We additionally enjoyed better-than-ever sightings of the tricky Barratt’s Warbler, aptly named Gorgeous Bushshrike, Crested Guineafowl, and Eastern Nicator to just name a few. Any trip to South Africa would be incomplete without mammals and our tally of 60 species included such difficult animals as the Aardvark, Aardwolf, Southern African Hedgehog, Bat-eared Fox, Smith’s Red Rock Hare and both Sable and Roan Antelopes. This really was a trip like no other! ____________________________________________________________________________________ Tour in Detail Our first full day of the tour began with a short walk through the gardens of our quaint guesthouse in Johannesburg. Here we enjoyed sightings of the delightful Red-headed Finch, small numbers of Southern Red Bishops including several males that were busy moulting into their summer breeding plumage, the near-endemic Karoo Thrush, Cape White-eye, Grey-headed Gull, Hadada Ibis, Southern Masked Weaver, Speckled Mousebird, African Palm Swift and the Laughing, Ring-necked and Red-eyed Doves. -

Status and Distribution of Faunal Diversity in Kafa Afromontane Coffee Forest
Status and Distribution of Faunal Diversity in Kafa Afromontane Coffee Forest Leykun Abunie Berhan Submitted to PPP Project July 2008 Addis Ababa Contents Executive Summary .....................................................................................................................4 Introduction..................................................................................................................................6 Literature Review Related to Faunal Diversity and Management...............................................8 Macro Policies and Priorities......................................................................................................8 Environmental Protection Policy.................................................................................................8 Wildlife Development / Management Policy................................................................................9 Analysis of Wildlife Sector in Ethiopia ......................................................................................10 Physical and Ecological Description of the Study Area ............................................................14 Objective of the Present Study...................................................................................................16 Methodology ..............................................................................................................................17 General Approach......................................................................................................................17 -

Development and Application of Survey Methods to Determine
Development and application of survey methods to determine habitat use in relation to forest management and habitat characteristics of the endangered numbat (Myrmecobius fasciatus) in the Upper Warren region, Western Australia by Anke Seidlitz BSc (Hons) A thesis submitted to Murdoch University to fulfil the requirements for the degree of Doctor of Philosophy in the discipline of Environmental and Conservation Sciences January 2021 Author’s declaration I declare that (a) the thesis is my own account of my research, except where other sources are acknowledged, (b) all co-authors, where stated and certified by my principal supervisor or executive author, have agreed that the works presented in this thesis represent substantial contributions from myself and (c) the thesis contains as its main content work that has not been previously submitted for a degree at any other university. ……………………………. Anke Seidlitz i Abstract Effective detection methods and knowledge on habitat requirements is key for successful wildlife monitoring and management. The numbat (Myrmecobius fasciatus) is an endangered, Australian-endemic marsupial that has experienced major population declines since European settlement. The Upper Warren region (UWR) in south-western Australia contains one of the two remaining natural populations. A lack of effective survey methods has caused a paucity of information regarding this population. This PhD project aimed to develop robust survey methods and determine habitat requirements for the numbat in the UWR. Given the perceived advantages of camera trap technology in wildlife research, camera trap trials were conducted to optimise camera methodologies for numbat detection. Swift 3C wide-angle camera traps positioned at ~25 cm above ground increased numbat detections by 140% compared to commonly used Reconyx PC900 camera traps. -
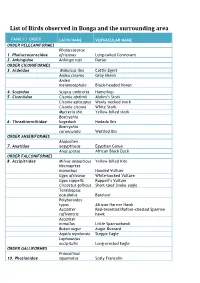
Birds Species in Bonga List
List of Birds observed in Bonga and the surrounding area FAMILY / ORDER LATIN NAME VERNACULAR NAME ORDER PELECANIFORMES Phalacrocorax 1. Phalacrocoracidae africanus Long-tailed Cormorant 2. Anhingidae Anhinga rufa Darter ORDER CICONIIFORMES 3. Ardeidae Bubulcus ibis Cattle Egret Ardea cinerea Grey Heron Ardea melanocephala Black-headed Heron 4. Scopidae Scopus umbretta Hamerkop 5. Ciconiidae Ciconia abdimii Abdim’s Stork Ciconia episcopus Wooly -necked stork Ciconia ciconia White Stork Mycteria ibis Yellow -billed stork Bostrychia 6. Threskiornithidae hagedash Hadada Ibis Bostrychia carunculata Wattled Ibis ORDER ANSERIFORMES Alopochen 7. Anatidae aegyptiacus Egyptian Goose Anas sparsa African Black Duck ORDER FALCONIFORMES 8. Accipitridae Milvus aegypticus Yellow -billed Kite Necrosyrtes monachus Hooded Vulture Gyps africanus White -backed Vulture Gyps ruppellii Ruppell’s Vulture Circaetus gallicus Short -toed Snake -eagle Terathopius ecaudatus Bateleur Polyboroides typus African Harrier Hawk Accipiter Red -breasted/Rufous -chested Sparrow rufiventris hawk Accipiter minullus Little Sparrowhawk Buteo augur Augur Buzzard Aquila nipalensis Steppe Eagle Lophoaetus occipitalis Long-crested Eagle ORDER GALLIFORMES Francolinus 10. Phasianidae squamatus Scaly Francolin FAMILY / ORDER LATIN NAME VERNACULAR NAME Francolinus castaneicollis Chestnut-napped Francolin ORDER GRUIFORMES Balearica 11. Gruidae pavonina Black Crowned Crane Bugeranus carunculatus Wattled crane Ruogetius 12. Rallidae rougetii Rouget’s Rail Podica 13. Heliornithidae senegalensis African Finfoot ORDER CHARADRIIFORMES 14. Scolopacidae Tringa ochropus Green Sandpiper Tringa hypolucos Common Sandpiper ORDER COLUMBIFORMES 15. Columbidae Columba guinea Speckled Pigeon Columba arquatrix African Olive Pigeon (Rameron Pigeon) Streptopelia lugens Dusky (Pink-breasted) Turtle Dove Streptopelia semitorquata Red-eyed Dove Turtur chalcospilos Emerald-spotted Wood Dove Turtur tympanistria Tambourine Dove ORDER CUCULIFORMES 17. Musophagidae Tauraco leucotis White -cheeked Turaco Centropus 18.