(Poaceae) in Iran
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
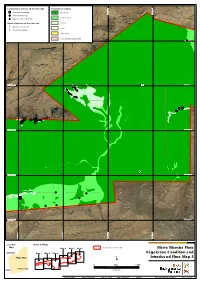
Additional Information
Current Survey Introduced Flora Records Vegetation Condition *Acetosa vesicaria Excellent 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 -

Review and Updated Checklist of Freshwater Fishes of Iran: Taxonomy, Distribution and Conservation Status
Iran. J. Ichthyol. (March 2017), 4(Suppl. 1): 1–114 Received: October 18, 2016 © 2017 Iranian Society of Ichthyology Accepted: February 30, 2017 P-ISSN: 2383-1561; E-ISSN: 2383-0964 doi: 10.7508/iji.2017 http://www.ijichthyol.org Review and updated checklist of freshwater fishes of Iran: Taxonomy, distribution and conservation status Hamid Reza ESMAEILI1*, Hamidreza MEHRABAN1, Keivan ABBASI2, Yazdan KEIVANY3, Brian W. COAD4 1Ichthyology and Molecular Systematics Research Laboratory, Zoology Section, Department of Biology, College of Sciences, Shiraz University, Shiraz, Iran 2Inland Waters Aquaculture Research Center. Iranian Fisheries Sciences Research Institute. Agricultural Research, Education and Extension Organization, Bandar Anzali, Iran 3Department of Natural Resources (Fisheries Division), Isfahan University of Technology, Isfahan 84156-83111, Iran 4Canadian Museum of Nature, Ottawa, Ontario, K1P 6P4 Canada *Email: [email protected] Abstract: This checklist aims to reviews and summarize the results of the systematic and zoogeographical research on the Iranian inland ichthyofauna that has been carried out for more than 200 years. Since the work of J.J. Heckel (1846-1849), the number of valid species has increased significantly and the systematic status of many of the species has changed, and reorganization and updating of the published information has become essential. Here we take the opportunity to provide a new and updated checklist of freshwater fishes of Iran based on literature and taxon occurrence data obtained from natural history and new fish collections. This article lists 288 species in 107 genera, 28 families, 22 orders and 3 classes reported from different Iranian basins. However, presence of 23 reported species in Iranian waters needs confirmation by specimens. -

Petrology and the Origin of the Intrusive Masses of the East of Jiroft
Revista Geoaraguaia ISSN:2236-9716 Barra do Garças - MT v.9, n.2, p.22-36. Agosto 2019 PETROLOGY AND THE ORIGIN OF THE INTRUSIVE MASSES OF THE EAST OF JIROFT PETROLOGIA E ORIGEM DE MATERIAIS INTRUSIVOS NO LESTE DE JIROFT Farbod Faraji Department of Geology, North Tehran Branch, Islamic Azad University, Tehran, Iran Afshin Ashja-Ardalan Department of Geology, North Tehran Branch, Islamic Azad University, Tehran, Iran [email protected] Moosa Kalimi-Noghreeian Department of Geology, Isfahan University, Isfahan, Iran Hamidreza Jafari Department of Geology, Sirjan Branch, Islamic Azad University, Sirjan, Iran ABSTRACT The study area with an area of 55 km2 is located south east of Kerman province and in the area of Jiroft city. This area is structurally and geological division in the Urmia-Dokhtar zone. The Urmia- Dokhtar volcanic belt part of the Alpine-Himalayan is a volcanic belt. Several intrusive bodies are in the east of Jiroft, which is part of Jebalbarez Batolite andigneous actives of JabalBarez area have occurred in four stages. The third magma activity of the region occurred in Oligomiocene and occurred during three phases. its lithological composition includes synogranite and Monzogranite, granodiorite, diorite, quartz monzonite. Quartz, Plagioclase and Potassium feldspar are the major minerals in granites. Biotite, Amphibole, espen, opac minerals are other manufactores of these rocks. Various types of granular, myrmekitic, Graphic and perthite textures are observed in them. Regarding field studies, petrographic, and geochemical studies, granite rocks of meta-aluminum and granitoid components of Iseries are volcanic arc of the continental margin of orogenic region that originate from melting of shell-shaped igneous rocks. -

192-210, 2011 Issn 1816-9112
192 Research Journal of Fisheries and Hydrobiology, 6(3): 192-210, 2011 ISSN 1816-9112 ORIGINAL ARTICLES The Probability Of Zagros Mountains Environmental Pollution Due To Seismic Response Of Bakhtiari Dam Zaniar Tokmechi Department of Civil Engineering, Mahabad Branch, Islamic Azad University, Mahabad, Iran. ABSTRACT The Bakhtiari Dam is a planned arch dam on the Bakhtiari River within the Zagros Mountains in Lorestan Province, Iran. At a planned height of 315 meters (1,033 ft), it will be the world's tallest dam once completed and withhold the largest reservoir in Iran. The Zagros Mountains are the largest mountain range in Iran and Iraq. With a total length of 1,500 km (932 mi), from northwestern Iran, and roughly correlating with Iran's western border, the Zagros range spans the whole length of the western and southwestern Iranian plateau and ends at the Straits of Hormuz. In this paper, the probability of environmental pollution due to heavy metals caused by Bakhtiari dam failure is studied. Finite Element and ZENGAR methods are used to analyze the probability of pollution at dam downstream. Different dam cross sections and various loading conditions are considered to study the effects of these factors on the seismic behavior of the dam. Results show that the effect of the highest cross section is not the most significant for heavy metals pollution at the dam down stream. Pollution coefficient due to stress along Y axis (Sy) is always the determinant pollution. While, in all sections Sx and Sy are the determinant parameter affecting downstream heavy metal pollution and normally are bigger than Sz. -

Wiregrass (Aristida Stricta)
Wiregrass (Aristida stricta) For definitions of botanical terms, visit en.wikipedia.org/wiki/Glossary_of_botanical_terms. Wiregrass is a perennial bunchgrass found in scrub, pinelands and coastal uplands throughout much of Florida. It is the dominant groundcover species in longleaf pine savannas and is a primary food source for gopher tortoises. Birds and small wildlife eat the seeds. Historically, cattle grazed on Wiregrass’s tender new growth. Wiregrass flowers are tiny and brown. They are born on spikelike terminal panicles Flower stalks are elongated and extend above the leaves. Leaf blades are long, thin and rolled inward, giving them a wiry appearance (hence the common name). They are erect and green when young Photo courtesy of Alan Cressler, Lady Bird Johnson Wildflower Center and begin to arch and turn brown as they age. Its fruits are small, yellowish caryopses. Seeds may be dispersed by wind, gravity or on the fur of passing animals. The genus name Aristida is from the Latin arista, meaning “awn” and referring to the three awns or bristle-like structures that extend from the florets. (An alternative common name is Pineland threeawn.) The species epithet, stricta, is from the Latin strictus, meaning straight or erect. Family: Poaceae (Grass family) Native range: Nearly throughout To see where natural populations of Wiregrass have been vouchered, visit www.florida.plantatlas.usf.edu. Hardiness: Zones 8A–10B Lifespan: Perennial Soil: Moist to dry, well-drained sandy soils Exposure: Full sun to partial shade Growth habit: 1–3’+ tall and equally wide Propagation: Division, seed Garden tips: Wiregrass is fast-growing and tolerant of drought conditions and low-nutrient soils. -

Asiatische Halictidae, 3. Die Artengruppe Der Lasioglossum Carinate-/Jvy/Ae«S (Insecta: Hymenoptera: Apoidea: Halictidae: Halictinae)
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at Linzer biol. Beitr. 27/2 525-652 29.12.1995 Asiatische Halictidae, 3. Die Artengruppe der Lasioglossum carinate-/jvy/ae«s (Insecta: Hymenoptera: Apoidea: Halictidae: Halictinae) A. W. EBMER Abstract: The palearctic species of the big species group Lasioglossum carinate- Evylaeus according to SAKAGAMI and other authors is presented. For the first time the numerous palearctic species are sorted into species groups. New taxa described are: Lasioglossum (Evylaeus) albipes villosum 9 (Rußland, Primorskij kraj), alexandrirtum 3 9 (Turkmenien, Kirgisien), eschaton 9 (Tadjikistan, Kirgisien, Usbekien, Turkmenien), anthrax 9 (China, Yunnan). Until recently unknown sexes of the following taxa are described for the first time: La- sioglossum {Evylaeus) reinigi EBMER 1978 3, nipponense (HIRASHIMA 1953) 3, salebrosum (BLÜTHGEN 1943) 9, apristum (VACHAL 1903) 3, epipygiale epipygiale (BLÜTHGEN 1924) 9, israelense EBMER 1974 3, calileps (BLÜTHGEN 1926) 9, sim- laense (CAMERON 1909) $, mesoviride EBMER 1974 3, leucopymatum (DALLA TORRE 1896) d\ edessae EBMER 1974 9, nursei (BLÜTHGEN 1926) 3, himalayense (BlNGHAM 1898) 3, perihirtulum (COCKERELL 1937) 3, suppressum EBMER 1983 9, oppositum (SMITH 1875) 3, funebre (CAMERON 1896) 9. Type specimens from A. Fedoenko's expedition to central Asia (1868-1871) have been studied and informations about localities are recorded which are not common knowledge to today entomologists. Einleitung Anlaß fur diese Publikation sind neue, umfangreichere Aufsammlungen vor allem aus Zentralasien. Schon vor vielen Jahren schenkte mir Dr. Wilhelm Grünwaldt, München, Aufsammlungen vor allem aus Turkmenien, aber auch aus den anderen, heute neuen Staaten Zentralasiens. Im Jahr 1990 schenkte mit Prof. Joachim Oehlke seine Auf- sammlungen aus Turkmenien sowie aus dem südlichen Kaukasus. -

Protocol Development for Somatic Embryogenesis, SSR Markers And
Asadi‑Aghbolaghi et al. Plant Methods (2021) 17:70 https://doi.org/10.1186/s13007‑021‑00768‑9 Plant Methods RESEARCH Open Access Protocol development for somatic embryogenesis, SSR markers and genetic modifcation of Stipagrostis pennata (Trin.) De Winter Masoumeh Asadi‑Aghbolaghi1, Beata Dedicova2* , Sonali Sachi Ranade2, Kim‑Cuong Le2, Farzad Sharifzadeh1, Mansoor Omidi1 and Ulrika Egertsdotter2 Abstract Background: Stipagrostis pennata (Trin.) De Winter is an important species for fxing sand in shifting and semi‑fxed sandy lands, for grazing, and potentially as a source of lignocellulose fbres for pulp and paper industry. The seeds have low viability, which limits uses for revegetation. Somatic embryogenesis ofers an alternative method for obtain‑ ing large numbers of plants from limited seed sources. Results: A protocol for plant regeneration from somatic embryos of S. pennata was developed. Somatic embryogen‑ esis was induced on Murashige & Skoog (MS) medium supplemented with 3 mg L–1 2,4‑D subsequently shoots were induced on MS medium and supplemented with 5 mg L–1 zeatin riboside. The highest· shoots induction was obtained when embryogenic callus derived from mature embryos· (96%) in combination with MS flter‑sterilized medium was used from Khuzestan location. The genetic stability of regenerated plants was analysed using ten simple sequence repeats (SSR) markers from S. pennata which showed no somaclonal variation in regenerated plants from somatic embryos of S. pennata. The regenerated plants of S. pennata showed genetic stability without any somaclonal varia‑ tion for the four pairs of primers that gave the expected amplicon sizes. This data seems very reliable as three of the PCR products belonged to the coding region of the genome. -

Phylogeny and Subfamilial Classification of the Grasses (Poaceae) Author(S): Grass Phylogeny Working Group, Nigel P
Phylogeny and Subfamilial Classification of the Grasses (Poaceae) Author(s): Grass Phylogeny Working Group, Nigel P. Barker, Lynn G. Clark, Jerrold I. Davis, Melvin R. Duvall, Gerald F. Guala, Catherine Hsiao, Elizabeth A. Kellogg, H. Peter Linder Source: Annals of the Missouri Botanical Garden, Vol. 88, No. 3 (Summer, 2001), pp. 373-457 Published by: Missouri Botanical Garden Press Stable URL: http://www.jstor.org/stable/3298585 Accessed: 06/10/2008 11:05 Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available at http://www.jstor.org/page/info/about/policies/terms.jsp. JSTOR's Terms and Conditions of Use provides, in part, that unless you have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and you may use content in the JSTOR archive only for your personal, non-commercial use. Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained at http://www.jstor.org/action/showPublisher?publisherCode=mobot. Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printed page of such transmission. JSTOR is a not-for-profit organization founded in 1995 to build trusted digital archives for scholarship. We work with the scholarly community to preserve their work and the materials they rely upon, and to build a common research platform that promotes the discovery and use of these resources. For more information about JSTOR, please contact [email protected]. -

Guidelines for Using the Checklist
Guidelines for using the checklist Cymbopogon excavatus (Hochst.) Stapf ex Burtt Davy N 9900720 Synonyms: Andropogon excavatus Hochst. 47 Common names: Breëblaarterpentyngras A; Broad-leaved turpentine grass E; Breitblättriges Pfeffergras G; dukwa, heng’ge, kamakama (-si) J Life form: perennial Abundance: uncommon to locally common Habitat: various Distribution: southern Africa Notes: said to smell of turpentine hence common name E2 Uses: used as a thatching grass E3 Cited specimen: Giess 3152 Reference: 37; 47 Botanical Name: The grasses are arranged in alphabetical or- Rukwangali R der according to the currently accepted botanical names. This Shishambyu Sh publication updates the list in Craven (1999). Silozi L Thimbukushu T Status: The following icons indicate the present known status of the grass in Namibia: Life form: This indicates if the plant is generally an annual or G Endemic—occurs only within the political boundaries of perennial and in certain cases whether the plant occurs in water Namibia. as a hydrophyte. = Near endemic—occurs in Namibia and immediate sur- rounding areas in neighbouring countries. Abundance: The frequency of occurrence according to her- N Endemic to southern Africa—occurs more widely within barium holdings of specimens at WIND and PRE is indicated political boundaries of southern Africa. here. 7 Naturalised—not indigenous, but growing naturally. < Cultivated. Habitat: The general environment in which the grasses are % Escapee—a grass that is not indigenous to Namibia and found, is indicated here according to Namibian records. This grows naturally under favourable conditions, but there are should be considered preliminary information because much usually only a few isolated individuals. -

Plants of Desert Dunes
Avinoam Danin Plants of Desert Dunes With 84 Figures Springer Contents Introduction Sand Deserts of the World . 2.1 Distribution of Sand Dunes 3 2.2 Sand Mobility 3 2.2.1 The Modes of Sand Movement 3 2.2.2 Ripples 5 2.2.3 Plants as Modifiers of Local Dune Topography 8 2.2.4 Microbiotic Crust as a Sand Fixer 10 2.2.4.1 The Succession of Sand Crust Community 11 2.2.4.2 Disturbances of the Microbiotic Crust 15 2.2.4.2.1 Natural Disturbances 15 2.2.4.2.2 Destruction by Humans and Domestic Animals 17 2.2.4.3 Distribution of Microbiotic Crust on Sands 17 Environmental Stresses in the Sand Desert 21 Substrate Stability 21 .1 Sand Accretion 21 .2 Deflation 22 .3 Blowouts 22 .4 Injuries by Airborne Sand 23 3.2 Moisture Regime 23 3.3 Nutrients 24 4 Habitat Types of Desert Dunes 25 4.1 Mobile Dunes 25 4.1.1 Crest 25 4.1.2 Slipface 25 4.1.3 Dune Base, Plinth 26 4.2 Stable Dunes 26 4.3 Sites of Constant Deflation 26 4.4 Sand Covering Other Substrates 27 4.4.1 Mobile Sand Covering Sand Sheets 27 4.4.2 Sand Covering Hills or Plains of Hard or Soft Rocks 27 4.4.3 Sand Covering Salt Marshes 27 VIII Contents. 4.4.4 Sand Covering Shallow Aquifers of Fresh Water 29 4.4.5 Sand Covering Dry Water Courses (Wadis Arroyos) 29 5 Plant Case Histories and Ecomorphological Types 31 5.1 Species Requiring Sand Accumulation - Stipagrostis scoparia Type 34 5.1.1 Stipagrostis scoparia 35 5.1.2 Stipagrostis acutiflora 38 5.1.3 Stipagrostis pungens 38 5.1.4 Stipagrostis pennata 39 5.1.5 Stipagrostis sabulicola and Other Southern African Species .. -

Department of the Interior Fish and Wildlife Service
Tuesday, February 10, 2009 Part II Department of the Interior Fish and Wildlife Service 50 CFR Part 17 Endangered and Threatened Wildlife and Plants; Determination of Endangered Status for Reticulated Flatwoods Salamander; Designation of Critical Habitat for Frosted Flatwoods Salamander and Reticulated Flatwoods Salamander; Final Rule VerDate Nov<24>2008 14:17 Feb 09, 2009 Jkt 217001 PO 00000 Frm 00001 Fmt 4717 Sfmt 4717 E:\FR\FM\10FER2.SGM 10FER2 erowe on PROD1PC63 with RULES_2 6700 Federal Register / Vol. 74, No. 26 / Tuesday, February 10, 2009 / Rules and Regulations DEPARTMENT OF THE INTERIOR during normal business hours, at U.S. Register on or before July 30, 2008, with Fish and Wildlife Service, Mississippi the final critical habitat rule to be Fish and Wildlife Service Fish and Wildlife Office, 6578 Dogwood submitted for publication in the Federal View Parkway, Jackson, MS 39213. Register by January 30, 2009. The 50 CFR Part 17 FOR FURTHER INFORMATION CONTACT: Ray revised proposed rule was signed on [FWS–R4–ES–2008–0082; MO 9921050083– Aycock, Field Supervisor, U.S. Fish and and delivered to the Federal Register on B2] Wildlife Service, Mississippi Field July 30, 2008, and it subsequently Office, 6578 Dogwood View Parkway, published on August 13, 2008 (73 FR RIN 1018–AU85 Jackson, MS 39213; telephone: 601– 47258). We also published supplemental information on the Endangered and Threatened Wildlife 321–1122; facsimile: 601–965–4340. If proposed rule to maintain the status of and Plants; Determination of you use a telecommunications device the frosted flatwoods salamander as Endangered Status for Reticulated for the deaf (TDD), call the Federal Information Relay Service (FIRS) at threatened (73 FR 54125; September 18, Flatwoods Salamander; Designation of 2008). -

Who's Related to Whom?
149 Who’s related to whom? Recent results from molecular systematic studies Elizabeth A Kellogg Similarities among model systems can lead to generalizations systematist’s question-why are there so many different about plants, but understanding the differences requires kinds of organisms? Studies of the evolution of develop- systematic data. Molecular phylogenetic analyses produce ment demand that the investigator go beyond the model results similar to traditional classifications in the grasses system and learn the pattern of variation in its relatives (Poaceae), and relationships among the cereal crops [3*]. This requires a reasonable assessment of the relatives’ are quite clear. Chloroplast-based phylogenies for the identity. Solanaceae show that tomato is best considered as a species of Solarium, closely related to potatoes. Traditional Knowledge of plant relationships has increased rapidly classifications in the Brassicaceae are misleading with in the past decade, reflecting partly the development regard to true phylogenetic relationships and data are only of molecular systematics. It has been known for some now beginning to clarify the situation. Molecular data are time that plant classifications do not reflect phylogeny also being used to revise our view of relationships among accurately, even though both phylogeny and classification flowering plant families. Phylogenetic data are critical for are hierarchical. The hierarchy of classification was interpreting hypotheses of the evolution of development. imposed in the late 18th century, well before ideas of descent with modification (evolution) were prevalent [4]. These pre-evolutionary groups were then re-interpreted in Address an evolutionary context, and were assumed to be products Harvard University Herbaria, 22 Divinity Avenue Cambridge, MA of evolution, rather than man-made artefacts.