Origin of High Stereocontrol in Olefin Cyclopropanation Catalyzed by An
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Supporting Information
Electronic Supplementary Material (ESI) for RSC Advances. This journal is © The Royal Society of Chemistry 2021 Supporting Information Synthesis, Oligomerization and Catalytic Studies of a Redox- Active Ni4-Cubane: A detailed Mechanistic Investigation Saroj Kumar Kushvaha, a† Maria Francis, b† Jayasree Kumar, a Ekta Nag, b Prathap Ravichandran, a b a Sudipta Roy* and Kartik Chandra Mondal* aDepartment of Chemistry, Indian Institute of Technology Madras, Chennai 600036, India. bDepartment of Chemistry, Indian Institute of Science Education and Research (IISER) Tirupati 517507, India. † Both authors contributed equally. Table of Contents 1. General remarks 2. Syntheses of complexes 1-3 3. Detail characterization of complexes 1-3 4. Computational details 5. X-ray single crystal diffraction of complexes 1-3. 6. Details of catalytic activities of complex 3 6.1. General procedures for the syntheses of aromatic heterocycles (6) and various diazoesters (7) 6.2. Optimization of reaction condition for cyclopropanation of aromatic heterocycles (6) 6.3. General synthetic procedure and characterization data for cyclopropanated aromatic heterocycles (8) 6.4. Determination of relative stereochemistry of 8 7. 1H and 13C NMR spectra of compounds 8a-j 8. Mechanistic investigations and mass spectrometric analysis 9. References S1 1. General remarks All catalytic reactions were performed under Argon atmosphere. The progress of all the catalytic reactions were monitored by thin layer chromatography (TLC, Merck silica gel 60 F 254) upon visualization of the TLC plate under UV light (250 nm). Different charring reagents, such as phosphomolybdic acid/ethanol, ninhydrin/acetic acid solution and iodine were used to visualize various starting materials and products spots on TLC plates. -

Gold-Catalyzed Ethylene Cyclopropanation
Gold-catalyzed ethylene cyclopropanation Silvia G. Rull, Andrea Olmos* and Pedro J. Pérez* Full Research Paper Open Access Address: Beilstein J. Org. Chem. 2019, 15, 67–71. Laboratorio de Catálisis Homogénea, Unidad Asociada al CSIC, doi:10.3762/bjoc.15.7 CIQSO-Centro de Investigación en Química Sostenible and Departamento de Química, Universidad de Huelva, Campus de El Received: 17 October 2018 Carmen 21007 Huelva, Spain Accepted: 11 December 2018 Published: 07 January 2019 Email: Andrea Olmos* - [email protected]; Pedro J. Pérez* - This article is part of the thematic issue "Cyclopropanes and [email protected] cyclopropenes: synthesis and applications". * Corresponding author Guest Editor: M. Tortosa Keywords: © 2019 Rull et al.; licensee Beilstein-Institut. carbene transfer; cyclopropane; cyclopropylcarboxylate; ethylene License and terms: see end of document. cyclopropanation; ethyl diazoacetate; gold catalysis Abstract Ethylene can be directly converted into ethyl 1-cyclopropylcarboxylate upon reaction with ethyl diazoacetate (N2CHCO2Et, EDA) F F in the presence of catalytic amounts of IPrAuCl/NaBAr 4 (IPr = 1,3-bis(2,6-diisopropylphenyl)imidazole-2-ylidene; BAr 4 = tetrakis(3,5-bis(trifluoromethyl)phenyl)borate). Introduction Nowadays the olefin cyclopropanation through metal-catalyzed carbene transfer starting from diazo compounds to give olefins constitutes a well-developed tool (Scheme 1a), with an exquisite control of chemo-, enantio- and/or diastereoselectiv- ity being achieved [1,2]. Previous developments have involved a large number of C=C-containing substrates but, to date, the methodology has not been yet employed with the simplest olefin, ethylene, for synthetic purposes [3]. Since diazo compounds bearing a carboxylate substituent are the most Scheme 1: (a) General metal-catalyzed olefin cyclopropanation reac- commonly employed carbene precursors toward olefin cyclo- tion with diazo compounds. -

Bond Distances and Bond Orders in Binuclear Metal Complexes of the First Row Transition Metals Titanium Through Zinc
Metal-Metal (MM) Bond Distances and Bond Orders in Binuclear Metal Complexes of the First Row Transition Metals Titanium Through Zinc Richard H. Duncan Lyngdoh*,a, Henry F. Schaefer III*,b and R. Bruce King*,b a Department of Chemistry, North-Eastern Hill University, Shillong 793022, India B Centre for Computational Quantum Chemistry, University of Georgia, Athens GA 30602 ABSTRACT: This survey of metal-metal (MM) bond distances in binuclear complexes of the first row 3d-block elements reviews experimental and computational research on a wide range of such systems. The metals surveyed are titanium, vanadium, chromium, manganese, iron, cobalt, nickel, copper, and zinc, representing the only comprehensive presentation of such results to date. Factors impacting MM bond lengths that are discussed here include (a) n+ the formal MM bond order, (b) size of the metal ion present in the bimetallic core (M2) , (c) the metal oxidation state, (d) effects of ligand basicity, coordination mode and number, and (e) steric effects of bulky ligands. Correlations between experimental and computational findings are examined wherever possible, often yielding good agreement for MM bond lengths. The formal bond order provides a key basis for assessing experimental and computationally derived MM bond lengths. The effects of change in the metal upon MM bond length ranges in binuclear complexes suggest trends for single, double, triple, and quadruple MM bonds which are related to the available information on metal atomic radii. It emerges that while specific factors for a limited range of complexes are found to have their expected impact in many cases, the assessment of the net effect of these factors is challenging. -

Catalytic Cyclopropanation of Polybutadienes
Erschienen in: Journal of Polymer Science, Part A: Polymer Chemistry ; 48 (2010), 20. - S. 4439-4444 https://dx.doi.org/10.1002/pola.24231 Catalytic Cyclopropanation of Polybutadienes JUAN URBANO,1 BRIGITTE KORTHALS,2 M. MAR DI´AZ-REQUEJO,1 PEDRO J. PE´ REZ,1 STEFAN MECKING2 1Laboratorio de Cata´ lisis Homoge´ nea, Departamento de Quı´mica y Ciencia de los Materiales, Unidad Asociada al CSIC, Centro de Investigacio´ n en Quı´mica Sostenible, Campus de El Carmen s/n, Universidad de Huelva, 21007 Huelva, Spain 2Department of Chemistry, University of Konstanz, 78464 Konstanz, Germany ABSTRACT: Catalytic cyclopropanation of commercial 1,2- or 1,4- bonyl-cyclopropene)]. Catalytic hydrogenation of residual dou- cis-polybutadiene, respectively, with ethyl diazoacetate catalyzed ble bonds of partially cyclopropanated polybutadienes provided by [TpBr3Cu(NCMe)] (TpBr3 ¼ hydrotris(3,4,5-tribromo-1-pyrazo- access to the corresponding saturated polyolefins. Thermal lyl)borate) at room temperature afforded high molecular weight properties are reported. 5 À1 (Mn > 10 mol ) side-chain or main-chain, respectively, carbox- yethyl cyclopropyl-substituted polymers with variable and con- trolled degrees of functionalization. Complete functionalization KEYWORDS: carbene addition; catalysis; functionalization of poly- of 1,4-cis-polybutadiene afforded poly[ethylene-alt-(3-ethoxycar- mers; organometallic catalysis; polar groups; polybutadienes INTRODUCTION Catalytic insertion polymerization of ethyl- polypropylene.6 Examples of catalytic post-polymerization ene and propylene is employed for the production of more reactions on saturated polyolefins are rare. The oxyfunction- than 60 million tons of polyolefins annually.1 These poly- alization of polyethylenes and polypropylenes by metal- mers are hydrocarbons, without any heteroatom-containing based catalysts can afford hydroxyl groups.7 We have functional groups, such as for example ester moieties. -
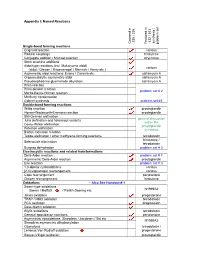
Appendix I: Named Reactions Single-Bond Forming Reactions Co
Appendix I: Named Reactions 235 / 335 432 / 533 synthesis / / synthesis Covered in Covered Featured in problem set problem Single-bond forming reactions Grignard reaction various Radical couplings hirstutene Conjugate addition / Michael reaction strychnine Stork enamine additions Aldol-type reactions (incl. Mukaiyama aldol) various (aldol / Claisen / Knoevenagel / Mannich / Henry etc.) Asymmetric aldol reactions: Evans / Carreira etc. saframycin A Organocatalytic asymmetric aldol saframycin A Pseudoephedrine glycinamide alkylation saframycin A Prins reaction Prins-pinacol reaction problem set # 2 Morita-Baylis-Hillman reaction McMurry condensation Gabriel synthesis problem set #3 Double-bond forming reactions Wittig reaction prostaglandin Horner-Wadsworth-Emmons reaction prostaglandin Still-Gennari olefination general discussion Julia olefination and heteroaryl variants within the Corey-Winter olefination prostaglandin Peterson olefination synthesis Barton extrusion reaction Tebbe olefination / other methylene-forming reactions tetrodotoxin hirstutene / Selenoxide elimination tetrodotoxin Burgess dehydration problem set # 3 Electrocyclic reactions and related transformations Diels-Alder reaction problem set # 1 Asymmetric Diels-Alder reaction prostaglandin Ene reaction problem set # 3 1,3-dipolar cycloadditions various [2,3] sigmatropic rearrangement various Cope rearrangement periplanone Claisen rearrangement hirstutene Oxidations – Also See Handout # 1 Swern-type oxidations (Swern / Moffatt / Parikh-Doering etc. N1999A2 Jones oxidation -

2 Results and Discussion
22 2 Results and Discussion 2 Results and Discussion 2.1 Syntheses of Azoninones - Unsaturated Nine-Membered Ring Lactams 2.1.1 Syntheses of Vinyl pyrrolidines N-Vinyl pyrrolidines are important reactants for the preparation of azoninones by zwitterionic aza- Claisen rearrangement. They can be obtained by derivatisation reactions starting from chiral molecules such as L-proline and 2S,4R-4-hydroxyproline or by metal catalysed cyclisation reactions of N- substituted allenic amines.66 Since large amounts of vinyl pyrrolidines have to be synthetically available for the systematic examination of the aza-Claisen rearrangement and its application in a total synthesis, especially the ex-chiral pool synthesis of these precursors has been extensively investigated by our group.67 The following two schemes represent the synthesis of the vinyl pyrrolidines [6] and [11] that have been used in this work as precursors for the aza-Claisen rearrangement. This synthesis allows the preparation of 50-100g of vinyl pyrrolidines. HO HO HO BnCl, Et3N TBSCl, imid. AcCl, MeOH CH Cl CH2Cl2 2 2 N COOH CO Me 100% N CO2Me 88% N 2 95% H H Bn [1] [2] [3] TBSO TBSO 1) Oxalyl chloride TBSO DMSO, Et3N, CH2Cl2 DIBALH, THF 2) Ph3P=CH2, THF N CO2Me N 89% N 70% Bn Bn OH Bn [4] [5] [6] Scheme 19 Synthesis of vinyl pyrrolidine [6] Allylamine [6] was efficiently generated via a six-step sequence starting from trans-4-hydroxy-L-(-)- proline [1] the overall yield was about 50% (Scheme 19). After esterification,68 the N-benzyl group 66 (a) Huby, N. -

Basic Research Needs for Catalysis Science
Basic Research Needs for Catalysis Science Report of the Basic Energy Sciences Workshop on Basic Research Needs for Catalysis Science to Transform Energy Technologies May 8–10, 2017 Image courtesy of Argonne National Laboratory. DISCLAIMER This report was prepared as an account of a workshop sponsored by the U.S. Department of Energy. Neither the United States Government nor any agency thereof, nor any of their employees or officers, makes any warranty, express or implied, or assumes any legal liability or responsibility for the accuracy, completeness, or usefulness of any information, apparatus, product, or process disclosed, or represents that its use would not infringe privately owned rights. Reference herein to any specific commercial product, process, or service by trade name, trademark, manufacturer, or otherwise, does not necessarily constitute or imply its endorsement, recommendation, or favoring by the United States Government or any agency thereof. The views and opinions of document authors expressed herein do not necessarily state or reflect those of the United States Government or any agency thereof. Copyrights to portions of this report (including graphics) are reserved by original copyright holders or their assignees, and are used by the Government’s license and by permission. Requests to use any images must be made to the provider identified in the image credits. This report is available in pdf format at https://science.energy.gov/bes/community-resources/reports/ REPORT OF THE BASIC RESEARCH NEEDS WORKSHOP FOR CATALYSIS SCIENCE Basic Research Needs for Catalysis Science TO TRANSFORM ENERGY TECHNOLOGIES Report from the U.S. Department of Energy, Office of Basic Energy Sciences Workshop May 8–10, 2017, in Gaithersburg, Maryland CHAIR: ASSOCIATE CHAIRS: Carl A. -

Cyclopropanation Catalyzed by Osmium Porphyrin Complexes Daniel A
Ames Laboratory Publications Ames Laboratory 3-1993 Cyclopropanation Catalyzed by Osmium Porphyrin Complexes Daniel A. Smith Iowa State University David N. Reynolds Iowa State University L. Keith Woo Iowa State University, [email protected] Follow this and additional works at: http://lib.dr.iastate.edu/ameslab_pubs Part of the Chemistry Commons The ompc lete bibliographic information for this item can be found at http://lib.dr.iastate.edu/ ameslab_pubs/362. For information on how to cite this item, please visit http://lib.dr.iastate.edu/ howtocite.html. This Article is brought to you for free and open access by the Ames Laboratory at Iowa State University Digital Repository. It has been accepted for inclusion in Ames Laboratory Publications by an authorized administrator of Iowa State University Digital Repository. For more information, please contact [email protected]. Cyclopropanation Catalyzed by Osmium Porphyrin Complexes Abstract Cyclopropanation of alkenes can be accomplished catalytically2 or stoichiometrically.3 Catalytic systems typically use a diazo reagent as the carbene source and a metal-containing mediator which forms a postulated metal carbene intermediate. Transfer of the carbene fragment from the metal to an alkene produces the cyclopropane product. Despite the wide variety of catalytic cyclopropanation systems, the putative carbene complex has never been isolated or observed in a catalytic system. This is somewhat surprising since the second category of cyclopropanation reactions involves the stoichiometric reaction -

Tandem-, Domino- and One-Pot Reactions Involving Wittig- And
DOI: 10.5772/intechopen.70364 Provisional chapter Chapter 1 Tandem-, Domino- and One-Pot Reactions Involving Wittig- and Horner-Wadsworth-Emmons Olefination Tandem-, Domino- and One-Pot Reactions Involving Wittig- and Horner-Wadsworth-Emmons Olefination Fatima Merza, Ahmed Taha and Thies Thiemann Fatima Merza, Ahmed Taha and Thies Thiemann Additional information is available at the end of the chapter Additional information is available at the end of the chapter http://dx.doi.org/10.5772/intechopen.70364 Abstract The Wittig olefination utilizing phosphoranes and the related Horner-Wadsworth- Emmons (HWE) reaction using phosphonates transform aldehydes and ketones into substituted alkenes. Because of the versatility of the reactions and the compatibility of many functional groups towards the transformations, both Wittig olefination and HWE reactions are a mainstay in the arsenal of organic synthesis. Here, an overview is given on Wittig- and Horner-Wadsworth-Emmons (HWE) reactions run in combination with other transformations in one-pot procedures. The focus lies on one-pot oxidation Wittig/HWE protocols, Wittig/HWE olefinations run in concert with metal catalyzed cross-coupling reactions, Domino Wittig/HWE—cycloaddition and Wittig-Michael transformations. Keywords: Wittig olefination, one-pot reactions, Domino reactions, tandem reactions, Horner-Wadsworth-Emmons olefination 1. Introduction The Wittig olefination utilizing phosphoranes and the related Horner-Wadsworth-Emmons (HWE) reaction using phosphonates transform aldehydes and ketones into substituted alkenes. Because of the versatility of the reactions and the compatibility of many functional groups in the transformations, both Wittig olefination and HWE reactions are a mainstay in the arsenal of organic synthesis. The mechanism of the Wittig olefination has been the subject of intense debate [1]. -

Cyclopropanation Chem 115
Myers Cyclopropanation Chem 115 Reviews: • Bonding Orbitals in Cyclopropane (Walsh Model): Roy, M.-N.; Lindsay, V. N. G.; Charette, A. B. Stereoselective Synthesis: Reactions of Carbon– Carbon Double Bonds (Science of Synthesis); de Vries, J. G., Ed.; Thieme: Stuttgart, 2011, Vol 1.; 731–817. Lebel, H.; Marcoux, J.-F.; Molinaro, C.; Charette, A. B. Chem. Rev. 2003, 103, 977–1050. Davies, H. M. L.; Beckwith, R. E. J. Chem. Rev. 2003, 103, 2861–2903. Li, A-H.; Dai, L. X.; Aggarwal, V. K. Chem. Rev. 1997, 97, 2341–2372. eS (") eA (") • Applications of Cyclopropanes in Synthesis Carson, C. A.; Kerr, M. A. Chem. Soc. Rev. 2009, 38, 3051–3060. Reissig, H.-U.; Zimmer, R. Chem. Rev. 2003, 103, 1151–1196. Gnad, F.; Reiser, O. Chem. Rev. 2003, 103, 1603–1624. • Cyclopropane Biosynthesis ! Thibodeaux, C. J.; Chang, W.-c.; Liu, H.-w. Chem. Rev. 2012, 112, 1681–1709. de Meijere, A. Angew. Chem. Int. Ed. 1979, 18, 809–886. General Strategies for Cyclopropanation: Introduction • via carbenoids "MCH2X" H HH H H H • via carbenes generated by decomposition of diazo compounds H H H H H H RCHN2 • Cyclopropanes are stable but highly strained compounds (ring strain ~29 kcal/mol). R • via Michael addition and ring closure • C–C bond angles = 60º (vs 109.5º for normal Csp3–Csp3 bonds). • Substituents on cyclopropanes are eclipsed. H–C–H angle is ~120º. As a result, the C–H bonds RCH–LG have higher s character compared to normal sp3 bonds. EWG EWG EWG LG R • Because of their inherent strain, the reactivity of cyclopropanes is more closely analogous to that of alkenes than that of alkanes. -

Synthesis of Functionalized Organic Molecules Using Copper Catalyzed
Duquesne University Duquesne Scholarship Collection Electronic Theses and Dissertations Fall 2011 Synthesis of Functionalized Organic Molecules Using Copper Catalyzed Cyclopropanation, Atom Transfer Radical Reactions and Sequential Azide- Alkyne Cycloaddition Carolynne Lacar Ricardo Follow this and additional works at: https://dsc.duq.edu/etd Recommended Citation Ricardo, C. (2011). Synthesis of Functionalized Organic Molecules Using Copper Catalyzed Cyclopropanation, Atom Transfer Radical Reactions and Sequential Azide-Alkyne Cycloaddition (Doctoral dissertation, Duquesne University). Retrieved from https://dsc.duq.edu/etd/1099 This Immediate Access is brought to you for free and open access by Duquesne Scholarship Collection. It has been accepted for inclusion in Electronic Theses and Dissertations by an authorized administrator of Duquesne Scholarship Collection. For more information, please contact [email protected]. SYNTHESIS OF FUNCTIONALIZED ORGANIC MOLECULES USING COPPER CATALYZED CYCLOPROPANATION, ATOM TRANSFER RADICAL REACTIONS AND SEQUENTIAL AZIDE-ALKYNE CYCLOADDITION A Dissertation Submitted to the Bayer School of Natural and Environmental Sciences Duquesne University In partial fulfillment of the requirements for the degree of Doctor of Philosophy By Carolynne Lacar Ricardo December 2011 Copyright by Carolynne Lacar Ricardo 2011 SYNTHESIS OF FUNCTIONALIZED ORGANIC MOLECULES USING COPPER CATALYZED CYCLOPROPANATION, ATOM TRANSFER RADICAL REACTIONS AND SEQUENTIAL AZIDE-ALKYNE CYCLOADDITION By Carolynne Lacar Ricardo Approved -

Recent Advances in the Iron-Catalyzed Cycloaddition Reactions
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Springer - Publisher Connector Review SPECIAL TOPIC July 2012 Vol.57 No.19: 23382351 Iron Catalysis in Synthetic Organic Chemistry doi: 10.1007/s11434-012-5141-z Recent advances in the iron-catalyzed cycloaddition reactions WANG ChunXiang & WAN BoShun* Dalian Institute of Chemical Physics, Chinese Academy of Sciences, Dalian 116023, China Received December 19, 2011; accepted February 13, 2012; published online April 26, 2012 The rapid generation of molecular in a relatively easy manner has made the cycloaddition reaction a powerful tool in the synthesis of different membered ring compounds. Clearly, iron catalysts are becoming a much more interesting and viable choice for this purpose. In this review, a number of promising results in the iron catalyzed cycloaddition reactions in the last decades were sum- marized. Special attention has been paid to the asymmetric cycloaddition reactions. cycloaddition, iron catalyst, asymmetric Citation: Wang C X, Wan B S. Recent advances in the iron-catalyzed cycloaddition reactions. Chin Sci Bull, 2012, 57: 23382351, doi: 10.1007/s11434-012-5141-z Cycloaddition reaction, which can construct several bonds in 2004, including addition, substitution, cycloaddition, re- simultaneously in a single step, is one of the most powerful duction and other reactions. At a later time, they also reviewed strategies for the synthesis of cyclic compounds from vari- the oxidative C–C coupling reactions [4] and the develop- ous unsaturated substrates such as alkynes, alkenes, allenes, ment of carbon-heteroatom and heteroatom-heteroatom bond nitriles, aldehydes, ketones, imines, etc.