Surfzone Hydrodynamics As a Key Determinant of Spatial Variation In
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Geomorphic Classification of Rivers
9.36 Geomorphic Classification of Rivers JM Buffington, U.S. Forest Service, Boise, ID, USA DR Montgomery, University of Washington, Seattle, WA, USA Published by Elsevier Inc. 9.36.1 Introduction 730 9.36.2 Purpose of Classification 730 9.36.3 Types of Channel Classification 731 9.36.3.1 Stream Order 731 9.36.3.2 Process Domains 732 9.36.3.3 Channel Pattern 732 9.36.3.4 Channel–Floodplain Interactions 735 9.36.3.5 Bed Material and Mobility 737 9.36.3.6 Channel Units 739 9.36.3.7 Hierarchical Classifications 739 9.36.3.8 Statistical Classifications 745 9.36.4 Use and Compatibility of Channel Classifications 745 9.36.5 The Rise and Fall of Classifications: Why Are Some Channel Classifications More Used Than Others? 747 9.36.6 Future Needs and Directions 753 9.36.6.1 Standardization and Sample Size 753 9.36.6.2 Remote Sensing 754 9.36.7 Conclusion 755 Acknowledgements 756 References 756 Appendix 762 9.36.1 Introduction 9.36.2 Purpose of Classification Over the last several decades, environmental legislation and a A basic tenet in geomorphology is that ‘form implies process.’As growing awareness of historical human disturbance to rivers such, numerous geomorphic classifications have been de- worldwide (Schumm, 1977; Collins et al., 2003; Surian and veloped for landscapes (Davis, 1899), hillslopes (Varnes, 1958), Rinaldi, 2003; Nilsson et al., 2005; Chin, 2006; Walter and and rivers (Section 9.36.3). The form–process paradigm is a Merritts, 2008) have fostered unprecedented collaboration potentially powerful tool for conducting quantitative geo- among scientists, land managers, and stakeholders to better morphic investigations. -

Animal Adaptations
Animal Adaptations The Animal Adaptations program at Hatfield Marine Science Center is designed to be a 50- minute lab-based program for 3rd-12th grade students that examines marine organisms from three different habitats (sandy beach, rocky shore and estuary) and explores the many ways they are adapted to their particular environment. This lab focuses on the adaptations of several groups of marine animals including Mollusks, Crustaceans, and Echinoderms, and investigates how they differ depending on whether they are found in a sandy beach, rocky shore, or estuary environment. Students will work in small groups with a variety of live animals, studying individual characteristics and how these organisms interact with their environment and one another. Background Information The Oregon Coast is made up of a series of rocky shores, sandy beaches, and estuaries, all of which are greatly affected by fluctuating tides. Many of these areas are intertidal and are alternately inundated by seawater and exposed to air, wind, and dramatic changes in temperature and salinity. High tide floods these areas with cold, nutrient laden seawater, bringing food to organisms that live there in the form of plankton and detritus. Low tides often expose these organisms to the dangers of predation and desiccation. In addition to tidal effects, organisms that inhabit sandy beaches and rocky shores also have to deal with the physical stresses of pounding waves. Because of these harsh conditions, organisms have developed special adaptations that not only help them to survive but thrive in these environments. An adaptation is a physical or behavioral trait that helps a plant or animal survive in a specific environment or habitat. -

Seashore the SANDYSEASHORE the Soilisinfertile, Anditisoften Windy, Andsalty
SEASHORE DESCRIPTION The seashore is an area fi lled with an interesting mix of unique plants and animals that have adapted to cope with this environment. Living on the edge of the sea is not easy. The soil is infertile, and it is often windy, dry and salty. THE SANDY SEASHORE Along a sandy shore there are no large rocks, algae or tidal pools. The sandy seashore can be divided into four general zones: Intertidal, Pioneer, Fixed Dune and Scrub Woodland. 1. Intertidal Zone: Between the low tide and the high tide mark is the intertidal zone. When the tide goes out the creatures living in this zone are left stranded. They have to endure the heat of the sun and the higher salinity of the water resulting from evaporation. Notice the many small holes on a sandy beach; they are the doorways to the homes of many animals which burrow under the sand where it is cooler. Some of Ecosystems of The Bahamas the creatures living in this zone are sea worms, sand fl eas and sand crabs. 2. Pioneer Zone: So named because it is where the fi rst plants to try to grow over sand. These plants must adapt to loose, shifting sand and poor soil. There is no protection from wind or salt spray. Plants here are usually low growing vines with waxy leaves. Some plants found in the pioneer zone are Purple seaside bean (C. rosea), Saltwort (Batis maritima), Goat's foot (Ipomea pes-caprae), and Sea purslane (Sesuvium portulacastrum). 3. Fixed Dune Zone: The next zone is the fi xed dune, so named because as the plants in the pioneer zone grip the sand around their roots and make the beach more stable, the sand mounds up into small humps or dunes. -

Morphologic Characteristics of the Blow River Delta, Yukon Territory, Canada
Louisiana State University LSU Digital Commons LSU Historical Dissertations and Theses Graduate School 1969 Morphologic Characteristics of the Blow River Delta, Yukon Territory, Canada. James Murl Mccloy Louisiana State University and Agricultural & Mechanical College Follow this and additional works at: https://digitalcommons.lsu.edu/gradschool_disstheses Recommended Citation Mccloy, James Murl, "Morphologic Characteristics of the Blow River Delta, Yukon Territory, Canada." (1969). LSU Historical Dissertations and Theses. 1605. https://digitalcommons.lsu.edu/gradschool_disstheses/1605 This Dissertation is brought to you for free and open access by the Graduate School at LSU Digital Commons. It has been accepted for inclusion in LSU Historical Dissertations and Theses by an authorized administrator of LSU Digital Commons. For more information, please contact [email protected]. This dissertation has been microfilmed exactly as received 70-252 McCLOY, James Murl, 1934- MORPHOLOGIC CHARACTERISTICS OF THE BLOW RIVER DELTA, YUKON TERRITORY, CANADA. The Louisiana State University and Agricultural and Mechanical College, Ph.D., 1969 Geography University Microfilms, Inc., Ann Arbor, Michigan Morphologic Characteristics of the Blow River Belta, Yukon Territory, Canada A Dissertation Submitted to the Graduate Faculty of the Louisiana State University and Agricultural and Mechanical College in partial fulfillment of the requirements for the degree of Doctor of Philosophy in The Department of Geography and Anthropology by James Murl McCloy B.A., State College at Los Angeles, 1961 May, 1969 ACKNOWLEDGEMENTS Research culminating in this dissertation was conducted under the auspices of the Arctic Institute of North America. The major portion of the financial support was received from the United States Army under contract no. BA-ARO-D-3I-I2I4.-G832, "Arctic Environmental Studies." Additional financial assistance during part of the writing stage was received in the form of a research assistantship from the Coastal Studies Institute, Louisi ana State University. -

PROTISTS Shore and the Waves Are Large, Often the Largest of a Storm Event, and with a Long Period
(seas), and these waves can mobilize boulders. During this phase of the storm the rapid changes in current direction caused by these large, short-period waves generate high accelerative forces, and it is these forces that ultimately can move even large boulders. Traditionally, most rocky-intertidal ecological stud- ies have been conducted on rocky platforms where the substrate is composed of stable basement rock. Projec- tiles tend to be uncommon in these types of habitats, and damage from projectiles is usually light. Perhaps for this reason the role of projectiles in intertidal ecology has received little attention. Boulder-fi eld intertidal zones are as common as, if not more common than, rock plat- forms. In boulder fi elds, projectiles are abundant, and the evidence of damage due to projectiles is obvious. Here projectiles may be one of the most important defi ning physical forces in the habitat. SEE ALSO THE FOLLOWING ARTICLES Geology, Coastal / Habitat Alteration / Hydrodynamic Forces / Wave Exposure FURTHER READING Carstens. T. 1968. Wave forces on boundaries and submerged bodies. Sarsia FIGURE 6 The intertidal zone on the north side of Cape Blanco, 34: 37–60. Oregon. The large, smooth boulders are made of serpentine, while Dayton, P. K. 1971. Competition, disturbance, and community organi- the surrounding rock from which the intertidal platform is formed zation: the provision and subsequent utilization of space in a rocky is sandstone. The smooth boulders are from a source outside the intertidal community. Ecological Monographs 45: 137–159. intertidal zone and were carried into the intertidal zone by waves. Levin, S. A., and R. -

Baja California Sur, Mexico)
Journal of Marine Science and Engineering Article Geomorphology of a Holocene Hurricane Deposit Eroded from Rhyolite Sea Cliffs on Ensenada Almeja (Baja California Sur, Mexico) Markes E. Johnson 1,* , Rigoberto Guardado-France 2, Erlend M. Johnson 3 and Jorge Ledesma-Vázquez 2 1 Geosciences Department, Williams College, Williamstown, MA 01267, USA 2 Facultad de Ciencias Marinas, Universidad Autónoma de Baja California, Ensenada 22800, Baja California, Mexico; [email protected] (R.G.-F.); [email protected] (J.L.-V.) 3 Anthropology Department, Tulane University, New Orleans, LA 70018, USA; [email protected] * Correspondence: [email protected]; Tel.: +1-413-597-2329 Received: 22 May 2019; Accepted: 20 June 2019; Published: 22 June 2019 Abstract: This work advances research on the role of hurricanes in degrading the rocky coastline within Mexico’s Gulf of California, most commonly formed by widespread igneous rocks. Under evaluation is a distinct coastal boulder bed (CBB) derived from banded rhyolite with boulders arrayed in a partial-ring configuration against one side of the headland on Ensenada Almeja (Clam Bay) north of Loreto. Preconditions related to the thickness of rhyolite flows and vertical fissures that intersect the flows at right angles along with the specific gravity of banded rhyolite delimit the size, shape and weight of boulders in the Almeja CBB. Mathematical formulae are applied to calculate the wave height generated by storm surge impacting the headland. The average weight of the 25 largest boulders from a transect nearest the bedrock source amounts to 1200 kg but only 30% of the sample is estimated to exceed a full metric ton in weight. -

Coastal Wetland Trends in the Narragansett Bay Estuary During the 20Th Century
u.s. Fish and Wildlife Service Co l\Ietland Trends In the Narragansett Bay Estuary During the 20th Century Coastal Wetland Trends in the Narragansett Bay Estuary During the 20th Century November 2004 A National Wetlands Inventory Cooperative Interagency Report Coastal Wetland Trends in the Narragansett Bay Estuary During the 20th Century Ralph W. Tiner1, Irene J. Huber2, Todd Nuerminger2, and Aimée L. Mandeville3 1U.S. Fish & Wildlife Service National Wetlands Inventory Program Northeast Region 300 Westgate Center Drive Hadley, MA 01035 2Natural Resources Assessment Group Department of Plant and Soil Sciences University of Massachusetts Stockbridge Hall Amherst, MA 01003 3Department of Natural Resources Science Environmental Data Center University of Rhode Island 1 Greenhouse Road, Room 105 Kingston, RI 02881 November 2004 National Wetlands Inventory Cooperative Interagency Report between U.S. Fish & Wildlife Service, University of Massachusetts-Amherst, University of Rhode Island, and Rhode Island Department of Environmental Management This report should be cited as: Tiner, R.W., I.J. Huber, T. Nuerminger, and A.L. Mandeville. 2004. Coastal Wetland Trends in the Narragansett Bay Estuary During the 20th Century. U.S. Fish and Wildlife Service, Northeast Region, Hadley, MA. In cooperation with the University of Massachusetts-Amherst and the University of Rhode Island. National Wetlands Inventory Cooperative Interagency Report. 37 pp. plus appendices. Table of Contents Page Introduction 1 Study Area 1 Methods 5 Data Compilation 5 Geospatial Database Construction and GIS Analysis 8 Results 9 Baywide 1996 Status 9 Coastal Wetlands and Waters 9 500-foot Buffer Zone 9 Baywide Trends 1951/2 to 1996 15 Coastal Wetland Trends 15 500-foot Buffer Zone Around Coastal Wetlands 15 Trends for Pilot Study Areas 25 Conclusions 35 Acknowledgments 36 References 37 Appendices A. -

New York State Artificial Reef Plan and Generic Environmental Impact
TABLE OF CONTENTS EXECUTIVE SUMMARY ...................... vi 1. INTRODUCTION .......................1 2. MANAGEMENT ENVIRONMENT ..................4 2.1. HISTORICAL PERSPECTIVE. ..............4 2.2. LOCATION. .....................7 2.3. NATURAL RESOURCES. .................7 2.3.1 Physical Characteristics. ..........7 2.3.2 Living Resources. ............. 11 2.4. HUMAN RESOURCES. ................. 14 2.4.1 Fisheries. ................. 14 2.4.2 Archaeological Resources. ......... 17 2.4.3 Sand and Gravel Mining. .......... 18 2.4.4 Marine Disposal of Waste. ......... 18 2.4.5 Navigation. ................ 18 2.5. ARTIFICIAL REEF RESOURCES. ............ 20 3. GOALS AND OBJECTIVES .................. 26 3.1 GOALS ....................... 26 3.2 OBJECTIVES .................... 26 4. POLICY ......................... 28 4.1 PROGRAM ADMINISTRATION .............. 28 4.1.1 Permits. .................. 29 4.1.2 Materials Donations and Acquisitions. ... 31 4.1.3 Citizen Participation. ........... 33 4.1.4 Liability. ................. 35 4.1.5 Intra/Interagency Coordination. ...... 36 4.1.6 Program Costs and Funding. ......... 38 4.1.7 Research. ................. 40 4.2 DEVELOPMENT GUIDELINES .............. 44 4.2.1 Siting. .................. 44 4.2.2 Materials. ................. 55 4.2.3 Design. .................. 63 4.3 MANAGEMENT .................... 70 4.3.1 Monitoring. ................ 70 4.3.2 Maintenance. ................ 72 4.3.3 Reefs in the Exclusive Economic Zone. ... 74 4.3.4 Special Management Concerns. ........ 76 4.3.41 Estuarine reefs. ........... 76 4.3.42 Mitigation. ............. 77 4.3.43 Fish aggregating devices. ...... 80 i 4.3.44 User group conflicts. ........ 82 4.3.45 Illegal and destructive practices. .. 85 4.4 PLAN REVIEW .................... 88 5. ACTIONS ........................ 89 5.1 ADMINISTRATION .................. 89 5.2 RESEARCH ..................... 89 5.3 DEVELOPMENT .................... 91 5.4 MANAGEMENT .................... 96 6. ENVIRONMENTAL IMPACTS ................. 97 6.1 ECOSYSTEM IMPACTS. -

Marine Biological Investigations in the Bahamas 23. Description of the Littoral Zonation at Nine Bahamian Rocky-Shore Localities
Marine biological investigations in the Bahamas 23. Description of the littoral zonation at nine Bahamian rocky-shore localities Hans Brattström Brattström H. 1999. Description of the littoral zonation at nine Bahamian rocky-shore localities. SARSIA Sarsia 84:319-365. Nine typical, more or less vertical, rocky-shore localities were visited in April-May 1967, and one of these was revisited in March 1968. The topographies of the stations are described and illustrated, and the vertical zonation of their flora and fauna is documented. On the whole the flora and fauna at the stations investigated was fairly poor. At least 37 species of algae and 85 animal species were recorded, but only 15 algae and 71 animals could be identified to species. Most of the algae were too undeveloped or grazed down to permit a reliable identification. Only half as many species were found at these nine stations as were found on three almost horizontal beach-rock stations described in an earlier article. Whereas many of the latter lived in a system of cavities below the rocks, all species found on the nine vertical stations were exposed to waves and the hot midday sun. The common yellow, black, grey, and white zones typical of the Bahamas, were often well-devel- oped. Some algae and many of the animals formed distinct belts within these zones, the higher up the more exposed the rocks were. Though tides and exposure presumably are responsible for the general zonation pattern, it is likely that in the cirripeds and vermetids the way of food-intake determines their upper limit. -

Understanding the Temporal Dynamics of the Wandering Renous River, New Brunswick, Canada
Earth Surface Processes and Landforms EarthTemporal Surf. dynamicsProcess. Landforms of a wandering 30, 1227–1250 river (2005) 1227 Published online 23 June 2005 in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/esp.1196 Understanding the temporal dynamics of the wandering Renous River, New Brunswick, Canada Leif M. Burge1* and Michel F. Lapointe2 1 Department of Geography and Program in Planning, University of Toronto, 100 St. George Street, Toronto, Ontario, M5S 3G3, Canada 2 Department of Geography McGill University, 805 Sherbrooke Street West, Montreal, Quebec, H3A 2K6, Canada *Correspondence to: L. M. Burge, Abstract Department of Geography and Program in Planning, University Wandering rivers are composed of individual anabranches surrounding semi-permanent of Toronto, 100 St. George St., islands, linked by single channel reaches. Wandering rivers are important because they Toronto, M5S 3G3, Canada. provide habitat complexity for aquatic organisms, including salmonids. An anabranch cycle E-mail: [email protected] model was developed from previous literature and field observations to illustrate how anabranches within the wandering pattern change from single to multiple channels and vice versa over a number of decades. The model was used to investigate the temporal dynamics of a wandering river through historical case studies and channel characteristics from field data. The wandering Renous River, New Brunswick, was mapped from aerial photographs (1945, 1965, 1983 and 1999) to determine river pattern statistics and for historical analysis of case studies. Five case studies consisting of a stable single channel, newly formed anabranches, anabranches gaining stability following creation, stable anabranches, and an abandoning anabranch were investigated in detail. -

Appendix E Fish Habitat Utilization Literature Review
Appendix E Fish Habitat Utilization Literature Review East Kitsap Nearshore Assessment Appendix E Appendix E - Fish Habitat Utilization Literature Review Toft et al. (2007) City Shoreline Fish Distribution Study objective: study the abundance and behavior of juvenile salmon and other fishes among various marine shoreline habitat types near the city of Seattle. Focused on 5 types of shorelines - cobble beach - sand beach - riprap extending into the upper intertidal zone - deep riprap extending into the subtidal - edge of overwater structures Also examined stomach contents of juvenile salmon. Fish sampling methods included enclosure nets and snorkel surveys. Riprap often results in greater beach slopes and steep embankments which effectively reduces the intertidal zone. These characteristics in turn, result in habitat loss for juvenile flatfishes. At high intertidal habitats, more crab were encountered at cobble beaches compared to sand beaches and riprap. Demersal fishes (cottids) were found in greater numbers at riprap. Differences in species density between habitat type were more apparent for demersal fishes than for pelagic species. Densities of salmon were significantly different among habitat types. The highest densities occurred at the edge of overwater structures and in deep riprap. Deep riprap creates some structured habitat which seems to attract surfperches and gunnels. Threespine stickleback, tubesnout, and bay pipefish were also more prevalent at deep riprap than at cobble, sand, and high elevation riprap. Beamer et al. (2006) – Habitat and fish use of pocket estuaries in the Whidbey basin and north Skagit county bays, 2004 and 2005 Pocket estuaries are characterized as having more dilute marine water relative to the surrounding estuary. -
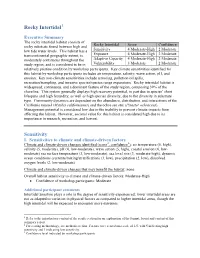
Rocky Intertidal Sensitivity
Rocky Intertidal1 Executive Summary The rocky intertidal habitat consists of Rocky Intertidal Score Confidence rocky substrate found between high and low tide water levels. This habitat has a Sensitivity 4 Moderate-High 2 Moderate transcontinental geographic extent, is Exposure 4 Moderate-High 2 Moderate moderately continuous throughout the Adaptive Capacity 4 Moderate-High 2 Moderate study region, and is considered to be in Vulnerability 3 Moderate 2 Moderate relatively pristine condition by workshop participants. Key climate sensitivities identified for this habitat by workshop participants includes air temperature, salinity, wave action, pH, and erosion. Key non-climate sensitivities include armoring, pollution/oil spills, recreation/trampling, and invasive species/species range expansions. Rocky intertidal habitat is widespread, continuous, and a dominant feature of the study region, composing 39% of the shoreline. This system generally displays high recovery potential, in part due to species’ short lifespans and high fecundity, as well as high species diversity, due to the diversity in substrate type. Community dynamics are dependent on the abundance, distribution, and interactions of the California mussel (Mytilus californianus) and the ochre sea star (Pisaster ochraceus). Management potential is considered low due to the inability to prevent climate impacts from affecting the habitat. However, societal value for this habitat is considered high due to its importance in research, recreation, and harvest. Sensitivity I. Sensitivities