<I>Cynobacteria Synechococcus</I> Sp. PCC 7002 Culture
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Chemical Resistance of Plastics
(c) Bürkle GmbH 2010 Important Important information The tables “Chemical resistance of plastics”, “Plastics and their properties” and “Viscosity of liquids" as well as the information about chemical resistance given in the particular product descriptions have been drawn up based on information provided by various raw material manufacturers. These values are based solely on laboratory tests with raw materials. Plastic components produced from these raw materials are frequently subject to influences that cannot be recognized in laboratory tests (temperature, pressure, material stress, effects of chemicals, construction features, etc.). For this reason the values given are only to be regarded as being guidelines. In critical cases it is essential that a test is carried out first. No legal claims can be derived from this information; nor do we accept any liability for it. A knowledge of the chemical and mechanical Copyright This table has been published and updated by Bürkle GmbH, D-79415 Bad Bellingen as a work of reference. This Copyright clause must not be removed. The table may be freely passed on and copied, provided that Extensions, additions and translations If your own experiences with materials and media could be used to extend this table then we would be pleased to receive any additional information. Please send an E-Mail to [email protected]. We would also like to receive translations into other languages. Please visit our website at http://www.buerkle.de from time to Thanks Our special thanks to Franz Kass ([email protected]), who has completed and extended these lists with great enthusiasm and his excellent specialist knowledge. -
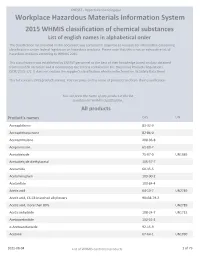
2015 WHMIS Classification : List of English Names in Alphabetical Order
CNESST - Répertoire toxicologique Workplace Hazardous Materials Information System 2015 WHMIS classification of chemical substances List of english names in alphabetical order The classification list provided in this document was compiled in response to requests for information concerning classifications under federal legislation on hazardous products. Please note that this is not an exhaustive list of hazardous products according to WHMIS 2015. This classification was established by CNESST personnel to the best of their knowledge based on data obtained from scientific literature and it incorporates the criteria contained in the Hazardous Products Regulations (SOR/2015-17). It does not replace the supplier's classification which can be found on its Safety Data Sheet. This list contains 2425 products names. You can press on the name of products to obtain their classification. You can press the name of any product in the list to obtain his WHMIS classification. All products Product's names CAS UN Acenaphthene 83-32-9 Acenaphthoquinone 82-86-0 Acenaphthylene 208-96-8 Acepromazine 61-00-7 Acetaldehyde 75-07-0 UN1089 Acetaldehyde diethylacetal 105-57-7 Acetamide 60-35-5 Acetaminophen 103-90-2 Acetanilide 103-84-4 Acetic acid 64-19-7 UN2789 Acetic acid, C6-C8 branched alkyl esters 90438-79-2 Acetic acid, more than 80% UN2789 Acetic anhydride 108-24-7 UN1715 Acetoacetanilide 102-01-2 o-Acetoacetaniside 92-15-9 Acetone 67-64-1 UN1090 2021-08-04 List of WHMIS controlled products 1 of 79 CNESST - Répertoire toxicologique Acetonitrile 75-05-8 UN1648 -

Résistance Chimique Des Plastiques », « Plastique Et Ses Propriétés » Et « Viscosité Des Liquides »
(c) Bürkle GmbH 2010 Importantes Informations Importantes Les tableaux « Résistance chimique des plastiques », « Plastique et ses propriétés » et « Viscosité des liquides ». ainsi que les informations au sujet de la résistance chimique donnée dans les descriptions de produit ont été élaborées à partir des informations fournies par différents fabricants de matières premières. Ces valeurs sont basées uniquement sur des essais en laboratoire avec des matières premières. Les composants en plastique déjà produisaient à partir de ces matières premières sont souvent soumis à des influences qui ne peuvent pas être reconnus dans les tests de laboratoire (température, pression, contrainte matérielle, les effets des produits chimiques, les caractéristiques de construction, etc.) Pour cette raison, les valeurs données doivent seulement être considérées comme des orientations. Dans les cas critiques, il est essentiel que le test soit effectué en premier. Aucune réclamation légal, ne peut pas être dérivé de cette information, ni nous n’avons pas la responsabilité pour cet-là. A la connaissance de la résistance chimique et mécanique seule n'est pas suffisante pour l'évaluation de l'utilisabilité d'un produit. Par exemple, la réglementation concernant les liquides inflammables (Prévention de l'explosion) doivent également être prises en considération. Droit d'auteur Ce tableau a été publié et mis à jour par Bürkle GmbH, D-79540 Lörrach comme un ouvrage de référence. Cette clause du droit d'auteur ne doit pas être enlevée. Le tableau peut être librement copié et transmis, à condition que les informations sur l'éditeur soient conservées. Extensions, les ajouts et les traductions Si vos propres expériences avec des matériaux et les médias pourraient être utilisés pour élargir cette table, alors nous serions heureux de recevoir toute information complémentaire. -

Risks of Iron Supplements S
Review Article Risks of iron supplements S. Umadevi*, A. Mohamed Akram, K. Mugesh, R. Mukilan ABSTRACT Iron is a vital mineral that is included in many over the counter multivitamin and mineral supplements and is used therapeutically in higher doses to treat or prevent iron deficiency anemia. When taken at the usual recommended daily doses or in replacement doses, iron has little or no adverse effect on the liver. However, in high doses or accidental overdoses, iron causes severe toxicities. Today different types of iron supplements are available in the market without mentioning the individual compound. These supplements are based on at least 20 different iron compounds and sold under a wide range of brands. Iron supplements are good for women and children, but when we focus on individual compound, not all supplements are created equal potency, as some compounds may actually do more harm than good. From the recent research, it was found that two iron compounds, ferric citrate and ferric ethylenediaminetetraacetic acid are often used in dietary supplements and as a food additive, respectively, in worldwide markets can increase the biomarkers for colon cancer, suggesting a link to the death toll. This review reports the cautious of iron supplements and serious disorders of an overdose of ionic compounds. KEY WORDS: Amphiregulin, Ferric citrate, Ferric ethylenediaminetetraacetic acid, Ferritin, Iron overload INTRODUCTION used, iron will enhance immunity, and it provides anticarcinogenic and cognition-enhancing activities.[1] Iron is a metallic element which is present in most of soils, mineral waters, and certain minerals. Iron The majority of the iron that is required for the body is an essential mineral that is compulsory for life. -

Workplace Hazardous Materials Information System
CNESST - Répertoire toxicologique Workplace Hazardous Materials Information System Classification of chemical substances List in CAS order with english name The classification list provided in this document was compiled in response to requests for information concerning classifications under federal legislation on controlled products. Please note that this is not an exhaustive list of hazardous products according to WHMIS 1988. This classification was established by CNESST personnel to the best of their knowledge based on data obtained from scientific literature and it incorporates the criteria contained in the Controlled Products Regulations (SOR/88-66). It does not replace the supplier’s classification which can be found on its Material Safety Data Sheet. You can press the name of any product in the list to obtain his WHMIS classification. All CAS numbers CAS Products' names 50-00-0 • Formaldehyde 50-21-5 • Lactic acid 50-23-7 • Dihydrocortisone 50-32-8 • Benzo(a)pyrene 50-78-2 • Acetylsalicylic acid 50-81-7 • Ascorbic acid 50-99-7 • Glucose 51-18-3 • Triethylnemelamine 51-28-5 • 2,4-Dinitrophenol 51-75-2 • N-Methyl-bis(2-chloroethyl)amine 51-78-5 • p-Aminophenol hydrochloride 51-79-6 • Urethane 52-89-1 • l-Cysteine hydrochloride 52-90-4 • l-Cysteine 53-70-3 • Dibenz(a,h)anthracene 53-96-3 • 2-Acetylaminofluorene 54-11-5 • Nicotine 54-21-7 • Sodium salicylate 54-64-8 • Ethylmercurithiosalicylic acid sodium salt 55-18-5 • N-Nitrosodiethylamine 55-55-0 • n-Methyl-p-aminophenol sulfate 55-68-5 • Phenylmercuric nitrate 56-03-1 • Biguanide