Influence of Cladophora Sp. on the Composition and Spatial Distribution of Macroinvertebrate Communities in Streams. a THESIS SU
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Susquehanna Riyer Drainage Basin
'M, General Hydrographic Water-Supply and Irrigation Paper No. 109 Series -j Investigations, 13 .N, Water Power, 9 DEPARTMENT OF THE INTERIOR UNITED STATES GEOLOGICAL SURVEY CHARLES D. WALCOTT, DIRECTOR HYDROGRAPHY OF THE SUSQUEHANNA RIYER DRAINAGE BASIN BY JOHN C. HOYT AND ROBERT H. ANDERSON WASHINGTON GOVERNMENT PRINTING OFFICE 1 9 0 5 CONTENTS. Page. Letter of transmittaL_.__.______.____.__..__.___._______.._.__..__..__... 7 Introduction......---..-.-..-.--.-.-----............_-........--._.----.- 9 Acknowledgments -..___.______.._.___.________________.____.___--_----.. 9 Description of drainage area......--..--..--.....-_....-....-....-....--.- 10 General features- -----_.____._.__..__._.___._..__-____.__-__---------- 10 Susquehanna River below West Branch ___...______-_--__.------_.--. 19 Susquehanna River above West Branch .............................. 21 West Branch ....................................................... 23 Navigation .--..........._-..........-....................-...---..-....- 24 Measurements of flow..................-.....-..-.---......-.-..---...... 25 Susquehanna River at Binghamton, N. Y_-..---...-.-...----.....-..- 25 Ghenango River at Binghamton, N. Y................................ 34 Susquehanna River at Wilkesbarre, Pa......_............-...----_--. 43 Susquehanna River at Danville, Pa..........._..................._... 56 West Branch at Williamsport, Pa .._.................--...--....- _ - - 67 West Branch at Allenwood, Pa.....-........-...-.._.---.---.-..-.-.. 84 Juniata River at Newport, Pa...-----......--....-...-....--..-..---.- -

Brook Trout Outcome Management Strategy
Brook Trout Outcome Management Strategy Introduction Brook Trout symbolize healthy waters because they rely on clean, cold stream habitat and are sensitive to rising stream temperatures, thereby serving as an aquatic version of a “canary in a coal mine”. Brook Trout are also highly prized by recreational anglers and have been designated as the state fish in many eastern states. They are an essential part of the headwater stream ecosystem, an important part of the upper watershed’s natural heritage and a valuable recreational resource. Land trusts in West Virginia, New York and Virginia have found that the possibility of restoring Brook Trout to local streams can act as a motivator for private landowners to take conservation actions, whether it is installing a fence that will exclude livestock from a waterway or putting their land under a conservation easement. The decline of Brook Trout serves as a warning about the health of local waterways and the lands draining to them. More than a century of declining Brook Trout populations has led to lost economic revenue and recreational fishing opportunities in the Bay’s headwaters. Chesapeake Bay Management Strategy: Brook Trout March 16, 2015 - DRAFT I. Goal, Outcome and Baseline This management strategy identifies approaches for achieving the following goal and outcome: Vital Habitats Goal: Restore, enhance and protect a network of land and water habitats to support fish and wildlife, and to afford other public benefits, including water quality, recreational uses and scenic value across the watershed. Brook Trout Outcome: Restore and sustain naturally reproducing Brook Trout populations in Chesapeake Bay headwater streams, with an eight percent increase in occupied habitat by 2025. -

Assessing Wetland Condition on a Watershed Basis in the Mid-Atlantic Region Using Synoptic Land-Cover Maps
ASSESSING WETLAND CONDITION ON A WATERSHED BASIS IN THE MID-ATLANTIC REGION USING SYNOPTIC LAND-COVER MAPS ROBERT P. BROOKS*, DENICE H. WARDROP, and JOSEPH A. BISHOP Penn State Cooperative Wetlands Center, 302 Walker Building, Pennsylvania State University, University Park, PA 16802 USA (*author for correspondence, phone: 814-863-1596, fax: 814-863-7943, e-mail:[email protected]) Abstract. We developed a series of tools to address three integrated tasks needed to effectively manage wetlands on a watershed basis: inventory, assessment, and restoration. Depending on the objectives of an assessment, availability of resources, and degree of confidence required in the results, there are three levels of effort available to address these three tasks. This paper describes the development and use of synoptic land-cover maps (Level 1) to assess wetland condition for a watershed. The other two levels are a rapid assessment using ground reconnaissance (Level 2) and intensive field assessment (Level 3). To illustrate the application of this method, seven watersheds in Pennsylvania were investigated representing a range of areas (89–777 km2), land uses, and ecoregions found in the Mid-Atlantic Region. Level 1 disturbance scores were based on land cover in 1-km radius circles centered on randomly-selected wetlands in each watershed. On a standardized, 100-point, human-disturbance scale, with 100 being severely degraded and 1 being the most ecologically intact, the range of scores for the seven watersheds was a relatively pristine score of 4 to a moderately degraded score of 66. This entire process can be conducted in a geographic information system (GIS)-capable office with readily available data and without engaging in extensive field investigations. -

2018 Pennsylvania Summary of Fishing Regulations and Laws PERMITS, MULTI-YEAR LICENSES, BUTTONS
2018PENNSYLVANIA FISHING SUMMARY Summary of Fishing Regulations and Laws 2018 Fishing License BUTTON WHAT’s NeW FOR 2018 l Addition to Panfish Enhancement Waters–page 15 l Changes to Misc. Regulations–page 16 l Changes to Stocked Trout Waters–pages 22-29 www.PaBestFishing.com Multi-Year Fishing Licenses–page 5 18 Southeastern Regular Opening Day 2 TROUT OPENERS Counties March 31 AND April 14 for Trout Statewide www.GoneFishingPa.com Use the following contacts for answers to your questions or better yet, go onlinePFBC to the LOCATION PFBC S/TABLE OF CONTENTS website (www.fishandboat.com) for a wealth of information about fishing and boating. THANK YOU FOR MORE INFORMATION: for the purchase STATE HEADQUARTERS CENTRE REGION OFFICE FISHING LICENSES: 1601 Elmerton Avenue 595 East Rolling Ridge Drive Phone: (877) 707-4085 of your fishing P.O. Box 67000 Bellefonte, PA 16823 Harrisburg, PA 17106-7000 Phone: (814) 359-5110 BOAT REGISTRATION/TITLING: license! Phone: (866) 262-8734 Phone: (717) 705-7800 Hours: 8:00 a.m. – 4:00 p.m. The mission of the Pennsylvania Hours: 8:00 a.m. – 4:00 p.m. Monday through Friday PUBLICATIONS: Fish and Boat Commission is to Monday through Friday BOATING SAFETY Phone: (717) 705-7835 protect, conserve, and enhance the PFBC WEBSITE: Commonwealth’s aquatic resources EDUCATION COURSES FOLLOW US: www.fishandboat.com Phone: (888) 723-4741 and provide fishing and boating www.fishandboat.com/socialmedia opportunities. REGION OFFICES: LAW ENFORCEMENT/EDUCATION Contents Contact Law Enforcement for information about regulations and fishing and boating opportunities. Contact Education for information about fishing and boating programs and boating safety education. -

West Branch Subbasin AMD Remediation Strategy
Publication 254 West Branch Susquehanna Subbasin May 2008 AMD Remediation Strategy: West Branch Susquehanna Background, Data Assessment River Task Force and Method Development Despite the enormous legacy ■ INTRODUCTION Pristine setting along the West Branch Susquehanna River. of pollution from abandoned mine The West Branch Susquehanna drainage (AMD) in the West Subbasin, draining a 6,978-square-mile Branch Susquehanna Subbasin, area in northcentral Pennsylvania, is the there has been mounting support largest of the six major subbasins in and enthusiasm for a fully restored the Susquehanna River Basin (Figure 1). watershed. Under the leadership The West Branch Susquehanna of Governor Edward G. Rendell Subbasin is one of extreme contrasts. While and with support from it has some of the Commonwealth’s Trout Unlimited, Pennsylvania most pristine and treasured waterways, Department of Environmental including 1,249 miles of Exceptional Protection Secretary Kathleen Value streams and scenic forestlands and mountains, it also unfortunately M. Smith McGinty established the West bears the legacy of past Branch Susquehanna River Task unregulated mining. With Abandoned mine lands in Clearfield County. Force (Task Force) in 2004. 1,205 miles of waterways The goal of the Task Force is to impaired by AMD, it is the assist and advise the department and most AMD-impaired region its partners as they work toward of the entire Susquehanna the long-term goal to remediate the River Basin (Figure 2). At its most degraded region’s AMD. sites, the West Branch The Task Force is comprised Susquehanna River contains of state, federal, and regional acidity concentrations of agencies, Trout Unlimited, and nearly 200 milligrams per other conservation and watershed liter (mg/l), and iron and aluminum concentrations of organizations (members are identified A. -

Northumberland County
NORTHUMBERLAND COUNTY START BRIDGE SD MILES PROGRAM IMPROVEMENT TYPE TITLE DESCRIPTION COST PERIOD COUNT COUNT IMPROVED Bridge replacement on Township Road 480 over Mahanoy Creek in West Cameron BASE Bridge Replacement Township Road 480 over Mahanoy Creek Township 3 $ 2,120,000 1 1 0 Bridge Replacement on State Route 1025 (Shakespeare Road) over Chillisquaque BASE Bridge Replacement State Route 1025 over Chillisquaque Creek Creek in East Chillisquaque Township, Northumberland County 1 $ 1,200,000 1 1 0 BASE Bridge Replacement State Route 4022 over Boile Run Bridge replacement on State Route 4022 over Boile Run in Lower Augusta Township 1 $ 195,000 1 0 0 Bridge replacement on State Route 2001 over Little Roaring Creek in Rush BASE Bridge Replacement State Route 2001 over Little Roaring Creek Township 1 $ 180,000 1 1 0 Bridge replacement on PA 405 over Norfolk Southern Railroad in West BASE Bridge Replacement PA 405 over Norfolk Southern Railroad Chillisquaque Township 1 $ 2,829,000 1 1 0 BASE Bridge Rehabilitation PA 61 over Shamokin Creek Bridge rehabilitation on PA 61 over Shamokin Creek in Coal Township 1 $ 850,000 1 0 0 Bridge rehabilitation on PA 45 over Chillisquaque Creek in East Chillisquaque & BASE Bridge Rehabilitation PA 45 over Chillisquaque Creek West Chillisquaque Townships 2 $ 1,700,000 1 0 0 Bridge replacement on State Route 2022 over Tributary to Shamokin Creek in BASE Bridge Replacement State Route 2022 over Tributary to Shamokin Creek Shamokin Township 3 $ 240,000 1 0 0 BASE Bridge Replacement Township Road 631 over -

Pennsylvania Code, Title 25, Chapter 93, Water Quality Standards
Presented below are water quality standards that are in effect for Clean Water Act purposes. EPA is posting these standards as a convenience to users and has made a reasonable effort to assure their accuracy. Additionally, EPA has made a reasonable effort to identify parts of the standards that are not approved, disapproved, or are otherwise not in effect for Clean Water Act purposes. Ch. 93 WATER QUALITY STANDARDS 25 CHAPTER 93. WATER QUALITY STANDARDS GENERAL PROVISIONS Sec. 93.1. Definitions. 93.2. Scope. 93.3. Protected water uses. 93.4. Statewide water uses. ANTIDEGRADATION REQUIREMENTS 93.4a. Antidegradation. 93.4b. Qualifying as High Quality or Exceptional Value Waters. 93.4c. Implementation of antidegradation requirements. 93.4d. Processing of petitions, evaluations and assessments to change a designated use. 93.5. [Reserved]. WATER QUALITY CRITERIA 93.6. General water quality criteria. 93.7. Specific water quality criteria. 93.8. [Reserved]. 93.8a. Toxic substances. 93.8b. Metals criteria. 93.8c. Human health and aquatic life criteria for toxic substances. 93.8d. Development of site-specific water quality criteria. 93.8e. Special criteria for the Great Lakes System. DESIGNATED WATER USES AND WATER QUALITY CRITERIA 93.9. Designated water uses and water quality criteria. 93.9a. Drainage List A. 93.9b. Drainage List B. 93.9c. Drainage List C. 93.9d. Drainage List D. 93.9e. Drainage List E. 93.9f. Drainage List F. 93.9g. Drainage List G. 93.9h. Drainage List H. 93.9i. Drainage List I. 93.9j. Drainage List J. 93.9k. Drainage List K. 93.9l. Drainage List L. -

Wild Trout Waters (Natural Reproduction) - September 2021
Pennsylvania Wild Trout Waters (Natural Reproduction) - September 2021 Length County of Mouth Water Trib To Wild Trout Limits Lower Limit Lat Lower Limit Lon (miles) Adams Birch Run Long Pine Run Reservoir Headwaters to Mouth 39.950279 -77.444443 3.82 Adams Hayes Run East Branch Antietam Creek Headwaters to Mouth 39.815808 -77.458243 2.18 Adams Hosack Run Conococheague Creek Headwaters to Mouth 39.914780 -77.467522 2.90 Adams Knob Run Birch Run Headwaters to Mouth 39.950970 -77.444183 1.82 Adams Latimore Creek Bermudian Creek Headwaters to Mouth 40.003613 -77.061386 7.00 Adams Little Marsh Creek Marsh Creek Headwaters dnst to T-315 39.842220 -77.372780 3.80 Adams Long Pine Run Conococheague Creek Headwaters to Long Pine Run Reservoir 39.942501 -77.455559 2.13 Adams Marsh Creek Out of State Headwaters dnst to SR0030 39.853802 -77.288300 11.12 Adams McDowells Run Carbaugh Run Headwaters to Mouth 39.876610 -77.448990 1.03 Adams Opossum Creek Conewago Creek Headwaters to Mouth 39.931667 -77.185555 12.10 Adams Stillhouse Run Conococheague Creek Headwaters to Mouth 39.915470 -77.467575 1.28 Adams Toms Creek Out of State Headwaters to Miney Branch 39.736532 -77.369041 8.95 Adams UNT to Little Marsh Creek (RM 4.86) Little Marsh Creek Headwaters to Orchard Road 39.876125 -77.384117 1.31 Allegheny Allegheny River Ohio River Headwater dnst to conf Reed Run 41.751389 -78.107498 21.80 Allegheny Kilbuck Run Ohio River Headwaters to UNT at RM 1.25 40.516388 -80.131668 5.17 Allegheny Little Sewickley Creek Ohio River Headwaters to Mouth 40.554253 -80.206802 -
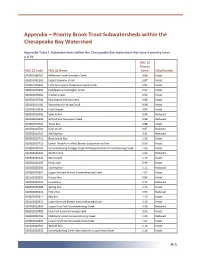
Appendix – Priority Brook Trout Subwatersheds Within the Chesapeake Bay Watershed
Appendix – Priority Brook Trout Subwatersheds within the Chesapeake Bay Watershed Appendix Table I. Subwatersheds within the Chesapeake Bay watershed that have a priority score ≥ 0.79. HUC 12 Priority HUC 12 Code HUC 12 Name Score Classification 020501060202 Millstone Creek-Schrader Creek 0.86 Intact 020501061302 Upper Bowman Creek 0.87 Intact 020501070401 Little Nescopeck Creek-Nescopeck Creek 0.83 Intact 020501070501 Headwaters Huntington Creek 0.97 Intact 020501070502 Kitchen Creek 0.92 Intact 020501070701 East Branch Fishing Creek 0.86 Intact 020501070702 West Branch Fishing Creek 0.98 Intact 020502010504 Cold Stream 0.89 Intact 020502010505 Sixmile Run 0.94 Reduced 020502010602 Gifford Run-Mosquito Creek 0.88 Reduced 020502010702 Trout Run 0.88 Intact 020502010704 Deer Creek 0.87 Reduced 020502010710 Sterling Run 0.91 Reduced 020502010711 Birch Island Run 1.24 Intact 020502010712 Lower Three Runs-West Branch Susquehanna River 0.99 Intact 020502020102 Sinnemahoning Portage Creek-Driftwood Branch Sinnemahoning Creek 1.03 Intact 020502020203 North Creek 1.06 Reduced 020502020204 West Creek 1.19 Intact 020502020205 Hunts Run 0.99 Intact 020502020206 Sterling Run 1.15 Reduced 020502020301 Upper Bennett Branch Sinnemahoning Creek 1.07 Intact 020502020302 Kersey Run 0.84 Intact 020502020303 Laurel Run 0.93 Reduced 020502020306 Spring Run 1.13 Intact 020502020310 Hicks Run 0.94 Reduced 020502020311 Mix Run 1.19 Intact 020502020312 Lower Bennett Branch Sinnemahoning Creek 1.13 Intact 020502020403 Upper First Fork Sinnemahoning Creek 0.96 -

TECHNIQUES for ESTIMATING MAGNITUDE and FREQUENCY of PEAK FLOWS for PENNSYLVANIA STREAMS by Marla H
U.S. Department of the Interior U.S. Geological Survey TECHNIQUES FOR ESTIMATING MAGNITUDE AND FREQUENCY OF PEAK FLOWS FOR PENNSYLVANIA STREAMS by Marla H. Stuckey and Lloyd A. Reed Water-Resources Investigations Report 00-4189 prepared in cooperation with PENNSYLVANIA DEPARTMENT OF TRANSPORTATION Lemoyne, Pennsylvania 2000 U.S. DEPARTMENT OF THE INTERIOR BRUCE BABBITT, Secretary U.S. GEOLOGICAL SURVEY Charles G. Groat, Director The use of product names in this report is for identification purposes only and does not constitute endorsement by the U.S. Government. For additional information Copies of this report may be write to: purchased from: District Chief U.S. Geological Survey U.S. Geological Survey Branch of Information Services 840 Market Street Box 25286 Lemoyne, Pennsylvania 17043-1586 Denver, Colorado 80225-0286 Email: [email protected] Telephone: 1-888-ASK-USGS ii CONTENTS Page Abstract . .1 Introduction . .1 Purpose and scope. .2 Previous investigations. .2 Development of flood-frequency prediction equations . .2 Description of stations used. .2 Basin characteristics used in equation development . .4 Regression analysis and resultant equations. .7 Limitations of regression equations . .12 Techniques for estimating magnitude and frequency of peak flows . .15 Summary and conclusions . .19 References cited . .19 Appendix 1. Basin characteristics for streamflow-gaging stations used in the development of the regional regression equations . .22 Appendix 2. Flood-flow frequencies computed from streamflow-gaging data and regression equations for streamflow-gaging stations used in analysis . .33 ILLUSTRATIONS Figures 1-3. Maps showing: 1. Streamflow-gaging stations used in development of flood-flow regression equations for Pennsylvania streams . 3 2. Carbonate regions in Pennsylvania. -

Download Report
2015 Associate Directors Dave Swank Retired Well Driller Since 1976 Staff Judy Becker Dr. Blair Carbaugh 2015 Directors District Manager Retired Professor, Lockhaven University Dave Crowl Former NCCD Board Member Chairman, Public Director Shirley Snyder Since 2006 Since 2006 Administrative Assistant Albert Mabus Leon Wertz Jaci Harner Manager of Geology/Environment, Vice‐Chairman, Farmer Director Watershed Specialist Eastern Industries, Inc. Since 2000 Former NCCD Board Member Since 2008 Michael McCleary Richard Shoch Erosion and Sediment Commissioner Director Technician Ted Carodiskey Since 2012 Little Shamokin Creek Watershed Association, Secretary Nathan Brophy Since 2010 John Kopp Agricultural Conservation Farmer Director Technician Since 2004 John Pfleegor Farmer Michael Erdley Former NCCD Board Member Since 2013 Public Director Since 2008 Michael Hubler Retired District Manager of the Dauphin Richard Daniels County Conservation District Farmer Director Since 2014 Since 2012 Gary Truckenmiller Farmer Director Since 2013 Left to right: Dave Crowl, Leon Wertz, Gary Truckenmiller, Richard Daniels, Michael Hubler, John Kopp, Richard Shoch EROSION AND SEDIMENTATION CONTROL PROGRAM The NCCD administers the Chapter 102 Erosion Control program through a signed delegation agreement with the Department of Environmental Protection (DEP). Under Chapter 102 delegation, NCCD conducts the following program responsibilities: community outreach, permit application receipt & review, permit approval or denial, consultation, site monitoring and final -

Columbia County Start Bridge Sd Program Improvement Type Title Description Cost Period Count Count
COLUMBIA COUNTY START BRIDGE SD PROGRAM IMPROVEMENT TYPE TITLE DESCRIPTION COST PERIOD COUNT COUNT BASE Bridge Replacement T-812 over Coles Creek Bridge replacement on T-812 over Coles Creek in Sugarloaf Township 2 $ 875,000 1 0 Bridge rehabilitation on State Route 4021 over Little Fishing Creek in Greenwood BASE Bridge Rehabilitation State Route 4021 over Little Fishing Creek Township 2 $ 1,200,000 1 0 Bridge replacement on State Route 1022 over Tributary to Green Creek in Fishing BASE Bridge Replacement State Route 1022 over Tributary to Green Creek Creek and Greenwood Townships 1 $ 140,000 1 1 Bridge replacement on State Route 1014 over Tributary to Little Briar Creek in Briar BASE Bridge Replacement State Route 1014 over Tributary to Little Briar Creek Creek Township 1 $ 150,000 1 1 Bridge replacement on PA 42 (Numidia Drive) over Roaring Creek in Conyngham BASE Bridge Replacement PA 42 over Roaring Creek Township 2 $ 1,400,000 1 1 Bridge replacement on State Route 44 (White Hall Road) over Chillisquaque Creek BASE Bridge Replacement SR 44 over Chillisquaque Creek in Madison Township, Columbia County. 1 $ 1,400,000 1 1 Bridge substructure rehabilitation on US 11 over Fishing Creek in the Town of BASE Bridge Rehabilitation US 11 over Fishing Creek Bloomsburg and Montour Township 3 $ 820,000 1 0 Bridge replacement on State Route 1033 over Little Pine Creek in Fishing Creek BASE Bridge Replacement State Route 1033 over Little Pine Creek Township 1 $ 235,000 1 0 Bridge replacement on State Route 2005 over Tributary to Roaring Creek in