Responses of Water Use Efficiency to Phenology in Typical Subtropical
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

US Universities in China Emphasize
United States Government Accountability Office Report to Congressional Requesters August 2016 CHINA U.S. Universities in China Emphasize Academic Freedom but Face Internet Censorship and Other Challenges Accessible Version GAO-16-757 August 2016 CHINA U.S. Universities in China Emphasize Academic Freedom but Face Internet Censorship and Other Challenges Highlights of GAO-16-757, a report to congressional requesters Why GAO Did This Study What GAO Found In its Country Reports on Human The 12 U.S. universities GAO reviewed generally reported receiving support for Rights Practices for 2015, the their institutions in China from Chinese government entities and universities, with Department of State (State) concluded limited funding from U.S. government agencies and other donors. Universities that academic freedom, a longstanding reported contributions from Chinese provincial and local governments and from concern in China, had recently partner universities for land, building construction, and use of campus facilities. worsened. At the same time, the Fewer than half of the universities reported receiving federal funding. Almost all number of U.S. universities of the U.S. universities said their programs in China generated net revenue for establishing degree-granting the university or had a neutral impact on its budget. institutions in partnership with Chinese universities—teaching predominantly Universities’ agreements with their Chinese partners or other policies that GAO Chinese students—has increased. reviewed generally include language protecting academic freedom or indicating While universities have noted that their institution in China would adhere to U.S. standards. About half of these institutions offer benefits, some universities GAO reviewed address access to information, such as providing academics and others have raised faculty and students with access to physical or online libraries, though few questions as to whether faculty, universities’ agreements and policies include language protecting Internet students, and staff may face restricted access. -

University Name Agency Number China Embassy in Tehran 3641
University Name Agency Number China Embassy in Tehran 3641 Aba Teachers College Agency Number 10646 Agricultural University of Hebei Agency Number 10086 Akzo vocational and technical College Agency Number 13093 Anglo-Chinese College Agency Number 12708 Anhui Agricultural University Agency Number 10364 Anhui Audit Vocational College Agency Number 13849 Anhui Broadcasting Movie And Television College Agency Number 13062 Anhui Business College of Vocational Technology Agency Number 12072 Anhui Business Vocational College Agency Number 13340 Anhui China-Australia Technology and Vocational College Agency Number 13341 Anhui College of Traditional Chinese Medicine Agency Number 10369 Anhui College of Traditional Chinese Medicine Agency Number 12924 Anhui Communications Vocational & Technical College Agency Number 12816 Anhui Eletrical Engineering Professional Technique College Agency Number 13336 Anhui Finance & Trade Vocational College Agency Number 13845 Anhui Foreign Language College Agency Number 13065 Anhui Industry Polytechnic Agency Number 13852 Anhui Institute of International Business Agency Number 13846 Anhui International Business and Economics College(AIBEC) Agency Number 12326 Anhui International Economy College Agency Number 14132 Anhui Lvhai Vocational College of Business Agency Number 14133 Anhui Medical College Agency Number 12925 Anhui Medical University Agency Number 10366 Anhui Normal University Agency Number 10370 Anhui Occupatinoal College of City Management Agency Number 13338 Anhui Police College Agency Number 13847 Anhui -

An Chengri an Chengri, Male, Born in November, 1964.Professor. Director
An Chengri , male, born in November, 1964.Professor. Director of Institute of International Studies, Department of Political Science, School of philosophy and Public Administration,Heilongjiang University. Ph. D student of Japanese politics and Diplomacy History, NanKai University,2001.Doctor(International Relations History), Kokugakuin University,2002. Research Orientation: Japanese Foreign Relations, International Relation History in East Asia Publications: Research on contemporary Japan-South Korea Relations(China Social Science Press,October,2008);International Relations History of East Asia(Jilin Science Literature Press,March,2005) Association: Executive Director of China Institute of Japanese History , Director of China Society of Sino-Japanese Relations History Address: No.74 Xuefu Road, Nangang District, Haerbin, Heilongjiang, Department of Political Science, School of philosophy and Public Administration,Heilongjiang University. Postcode: 150080 An shanhua , Female, born in July,1964. Associate Professor, School of History, Dalian University. Doctor( World History),Jilin University,2007. Research Orientation: Modern and contemporary Japanese History, Japanese Foreign Relations, Political Science Publications: Comparative Studies on World Order View of China Korea and Japan and their Diplomatic in Modern Time ( Japanese Studies Forum , Northeast Normal University, 2006); Analysis of Japan's anti-system ideology towards the international system ( Journal of Changchun University of Science and Technology , Changchun University,2006) -

Global Hospitality and Tourism Management Technologies
An Excellent Addition to Your Library! Released: August 2011 Global Hospitality and Tourism Management Technologies Patricia Ordóñez de Pablos (The University of Oviedo, Spain), Robert Tennyson (University of Minnesota, USA) and Jingyuan Zhao (Harbin Institute of Technologies, China) Tourism is a dynamic part of our economy. The global hospitality and tourism industry, which blends the lodging, food, attractions, cultural, and travel industries, is the world’s largest industry with $4.5 trillion in expenditures gener- ating 212 million jobs. Dynamic market forces such as global competition, changing customer expectations, and new communication technology—combined with career challenges due to ongoing industry consolidation and corporate restructuring—mandate continuous learning and the sharing of ideas. Global Hospitality and Tourism Management Technologies is a comprehensive collection which aims to be a source of information for all those interested in tourism and hospitality management, approaches, and trends, as well as to cover the emerging research topics which seek to define the future of IT and cultural development in the 21st century. The book provides a reference for policymakers, government officers, academics, and practitioners interested in understanding applications of IT for tourism and hospitality management. Topics Covered: • Beverage Management • Environmental Issues, Tourism and Leisure • Club Management • E-Tourism • Cultural Differences and Tourism Strategies • European Union Policies for Tourism • Eco-Tourism and Sustainable Tourism • Globalization and Tourism • Emerging Markets for Tourism • Green Management in Tourism and Hospitality ISBN: 9781613500415; © 2012; 270 pp. Print: US $185.00 | Perpetual: US $280.00 | Print + Perpetual: US $370.00 Market: This premier publication is essential for all academic and research library reference collections. -

A Complete Collection of Chinese Institutes and Universities For
Study in China——All China Universities All China Universities 2019.12 Please download WeChat app and follow our official account (scan QR code below or add WeChat ID: A15810086985), to start your application journey. Study in China——All China Universities Anhui 安徽 【www.studyinanhui.com】 1. Anhui University 安徽大学 http://ahu.admissions.cn 2. University of Science and Technology of China 中国科学技术大学 http://ustc.admissions.cn 3. Hefei University of Technology 合肥工业大学 http://hfut.admissions.cn 4. Anhui University of Technology 安徽工业大学 http://ahut.admissions.cn 5. Anhui University of Science and Technology 安徽理工大学 http://aust.admissions.cn 6. Anhui Engineering University 安徽工程大学 http://ahpu.admissions.cn 7. Anhui Agricultural University 安徽农业大学 http://ahau.admissions.cn 8. Anhui Medical University 安徽医科大学 http://ahmu.admissions.cn 9. Bengbu Medical College 蚌埠医学院 http://bbmc.admissions.cn 10. Wannan Medical College 皖南医学院 http://wnmc.admissions.cn 11. Anhui University of Chinese Medicine 安徽中医药大学 http://ahtcm.admissions.cn 12. Anhui Normal University 安徽师范大学 http://ahnu.admissions.cn 13. Fuyang Normal University 阜阳师范大学 http://fynu.admissions.cn 14. Anqing Teachers College 安庆师范大学 http://aqtc.admissions.cn 15. Huaibei Normal University 淮北师范大学 http://chnu.admissions.cn Please download WeChat app and follow our official account (scan QR code below or add WeChat ID: A15810086985), to start your application journey. Study in China——All China Universities 16. Huangshan University 黄山学院 http://hsu.admissions.cn 17. Western Anhui University 皖西学院 http://wxc.admissions.cn 18. Chuzhou University 滁州学院 http://chzu.admissions.cn 19. Anhui University of Finance & Economics 安徽财经大学 http://aufe.admissions.cn 20. Suzhou University 宿州学院 http://ahszu.admissions.cn 21. -

About the Contributors
262 About the Contributors Tomas Gabriel Bas earned his Ph.D. in Technology Management and Marketing of Innovation from University of Quebec (Canada). He has a M.Sc. in Environment from University of Montreal (Canada). He has two post-doctoral degrees in Technology Innovation Clusters from University of Quebec (Can- ada) and in Knowledge and Technology Innovation in Natural Resources Industries from Universidad Adolfo Ibañez (Chile). His research projects focus on biotechnology management, technological in- novation, clusters, knowledge in natural resources, and public policies. He is Professor of Technology and Innovation Management at the University Adolfo Ibáñez (Chile) and consultant for World Bank, United Nations, IDRC, Statistics Canada, and many multinationals firms. Honors include: “Who’s Who in the World,” 28th Edition – 2010 and 2011; “Top 100 Scientists 2011” International Biographical Centre, Cambridge, England; “2000 Outstanding Intellectuals of the 21st Century – 2011,” Interna- tional Biographical Centre, Cambridge, England; and “Bright Minds - Brain World Class.” He is one of the 10’s most influential researchers of Chile in Economics and Management (according to interna- tional citation index ISI). Jingyuan Zhao is an Associate Professor at School of Management, Beijing Union University, China. Her PhD is in Management Science and Engineering in Chinese Academy of Sciences & University of Science and Technology of China. She has worked on two Postdoctoral Programs in the Technol- ogy Economy and Management from Harbin Institute of Technology, China, and the Management of Technology from the Canada Research Chair, University of Québec at Montréal, Canada. Dr. Zhao’s expertise is on regional innovation management, high-tech industry cluster, knowledge management, technology diffusion, and organization learning. -

March 2018 Updated 3+2 Partnership List
March, 2018 UH Manoa 3+2 Partnership List 3+2 Partner Institutions Major Programs at UHM Agt Signed Program Start Entomology Tropical Plant and Soil Agricultural University of Hebei (HBAU) pending Sciences Tropical Plant Pathology Asian Studies March, 2015 Fall 2016 Beijing Foreign Studies University (BFSU) Second Language Studies July, 2016 Fall 2017 Communication June, 2016 Fall 2017 Beijing University of Civil Engineering and Architecture (BUCEA) Engineering Pending Central South University (CSU) Second Language Studies March, 2018 Fall 2018 Linguistics All Education Programs China University of Geosciences (CUGW) Geology & Geophysics May, 2016 Fall 2017 Fudan University (FDU) Public Health September, 2016 Fall 2017 Hainan Normal University (HNNU) All Education March, 2018 Fall 2018 Hainan University (HNU) All Agriculture Programs December, 2015 Fall 2016 Huaqiao University of Foreign Second Language Studies pending Languages (HQUFL) Education Huazhong Agricultural University Human Nutrition, Food & (HZAU) Animal Sciences September, 2016 Fall 2017 Hunan Agricultural University (HAU) All Agricultural Programs March, 2015 SPRING 2016 Nanchang University (NCU) Public Health July, 2015 Fall 2017 Nankai University (NKU) Economics April, 2015 Fall 2016 Northwest A&F University (NWAFU) All Agricultural Programs May, 2015 Fall 2016 Second Language Studies October, 2016 Fall 2017 Social Sciences October, 2016 Fall 2016 Nanjing University of Information SOEST - Atmospheric Science and Technology (UESTC) Sciences September, 2014 Fall 2015 Renmin -
Sorbus Lushanensis, a New Species of Rosaceae from China
A peer-reviewed open-access journal PhytoKeys 119: 97–105 (2019)Sorbus lushanensis, a new species of Rosaceae from China 97 doi: 10.3897/phytokeys.119.32148 RESEARCH ARTICLE http://phytokeys.pensoft.net Launched to accelerate biodiversity research Sorbus lushanensis, a new species of Rosaceae from China Jing Qiu1,2, Yang Zhao1, Qi Qi1, Xin Chen1 1 Co-Innovation Center for Sustainable Forestry in Southern China, College of Biology and The Environment, Nanjing Forestry University, Nanjing 210037, Jiangsu China 2 Sanjiang University, Nanjing 210012, Jiangsu, China Corresponding author: Xin Chen ([email protected]) Academic editor: Hanno Schaefer | Received 4 December 2018 | Accepted 10 March 2019 | Published 26 March 2019 Citation: Qiu J, Zhao Y, Qi Q, Chen X (2019) Sorbus lushanensis, a new species of Rosaceae from China. PhytoKeys 119: 97–105. https://doi.org/10.3897/phytokeys.119.32148 Abstract Sorbus lushanensis Xin Chen & Jing Qiu, sp. n. (Rosaceae), a new simple-leaved species belonging to Sorbus subg. Aria sect. Alnifoliae, is described from Anhui and Jiangxi provinces in China. Illustrations, photographs of wild plants and a distribution map are presented. The new species is morphologically similar to S. folgneri, but can be distinguished easily by its abaxially greenish-grey tomentose leaves, scale- like stipules and glabrous styles. Keywords Sorbus, new species, taxonomy, China Introduction Species of Sorbus are mainly distributed in the temperate areas of the Northern Hemi- sphere, with a centre of the highest diversity in East Asia, especially in China. The genus comprises about 100 (Lu and Spongberg 2003) to more than 250 (Phipps et al. -

Table of Contents
Table of Contents Preface .................................................................................................................................................xiii Chapter 1 Dynamic Competitive of Geographic Clusters (Such as Micro Regional Systems) in the Generation of Innovative Capabilities ....................................................................................................................... 1 Tomas Gabriel Bas, Universidad Adolfo Ibañez, Chile Chapter 2 Innovation by Imitation? Benchmarking Success Stories of Regional Innovation .............................. 14 Gert-Jan Hospers, University of Twente, The Netherlands & Radboud University, The Netherlands Erwin van Tuijl, Erasmus University Rotterdam, The Netherlands Paul Benneworth, University of Twente, The Netherlands Chapter 3 High Technology Cluster Growth Initiatives in Singapore ................................................................... 26 Pak Tee Ng, Nanyang Technological University, Singapore Chapter 4 Technology Policy Perspective: Critical Review of Software Technology Parks in India ................... 37 Neeta Baporikar, Ministry of Higher Education, CAS - Salalah, Sultanate of Oman Chapter 5 Canadian Biotechnology Systems: The Stakeholders and the Institutional Infrastructure ................... 53 Johanne Queenton, University of Sherbrooke, Canada Yvon Dufour, University of Sherbrooke, Canada Régis Milot, University of Sherbrooke, Canada Chapter 6 Creative Regions in Europe: Exploring Creative Industry Agglomeration and the Wealth -

A Critical Reading of Multiculturalism in English Language Teaching and Learning
Nov. 2008, Volume 5, No.11 (Serial No.48) US-China Education Review, ISSN1548-6613, USA A critical reading of multiculturalism in English language teaching and learning ZHAO Ning (English Department, Sanjiang University, Nanjing Jiangsu 210012, China) Abstract: Multiculturalism becomes the buzzword in educational fields. The implication of multiculturalism is critically examined to appreciate its relevance to English language teaching. Critical multiculturalism is advocated whereby L2 learners need to examine how social and historical discourses construct their identities and have a critical reflection about the discourses’ power/knowledge and social impact. Viewing language and culture as artificial systems with ideological dominance, critical multiculturalism leads to a critical pedagogy to negotiate, reverse and resist such ideological dominance. Critical multiculturalism enables L2 teachers and learners to create, maintain and transform language uses, hence their identities and social realities. Key words: critical multiculturalism; discourse, identity; power; language teaching 1. A critical reading of multiculturalism Globalization is evident in numerous spheres of our everyday life with persistent penetration of the global capital and increasing immigration from developing countries to developed countries, resulting in the phenomenal growth of the culturally diverse L2 learners. Globalization also brings destabilization and recreation of identities in Diaspora for L2 learners. Nowadays, multiculturalism becomes the buzzword in political and educational arenas. A critical examination of the implication of multiculturalism is needed to appreciate its relevance to English language teaching. The understanding of multiculturalism has to start with culture, discourse, identity and power. Culture refers to meanings embedded in social interactions, which involves two aspects—discourse and identity, both of them are social constructions. -

CURRICULUM VITAE Dr. GAO Yanping
CURRICULUM VITAE Dr. GAO Yanping Institute of Literature, Chinese Academy of Social Sciences, 5 Jianguomennei Street, Beijing, 100732, China Tel and Fax: 86-010-85195457; Mobile: 86- 18611339203 E-mail: [email protected] ; [email protected] [email protected] Personal Information: Date and place of birth: January 11, 1978: Ningbo, China Studies and Career: - 1995-1999: BA Studies in the History Department of Lanzhou University, China. - 1999-2002: MA in Literary Theory, Zhejiang University, China. Supervisor, prof. Wang Yu- anxiang. - 2003-2008: Lecturer in the Chinese Department of Sanjiang University (Nanjing), Taught courses in literature, literary theory, aesthetics and culture studies - 2006-2009 Editor responsible for introducing contemporary foreign literature and theoretical works for Nanjing University Press and Yilin Press. Served as the editor for translations of Eng- lish books for Nanjing University Press, 2006-2009, including A Companion to Cultural Studies (edited by Toby Miller), The Conquest of Cool: Business Culture, Counterculture, and the Rise of Hip Consumerism (by Thomas Frank), Einstein, History, and Other Passions: The Rebellion Against Science at The End of the Twentieth Century (by Gerald Holton ), etc. Served as the edi- tor responsible for literary translations for Yilin Press, 2002-2003, I edited books including: Bab- bitt (by S. Lewis), The Famished Road (by Ben Okri), The Human Stain (by P. Roth), etc. - 2008-2011, PhD-studies in Aesthetics, Philosophy Department of CASS (Chinese Academy of Social Sciences) Graduate School. Title:”Winckelmann’s Study of Greek Art,” Supervisor Prof. Wang Keping. Defense: May, 2011. - From 2011 till present, Assistant Research Fellow (parallel to Assistant Professor, with tenure), Theory Department of the Institute of Literature, CASS, involved in Aesthetics and Art studies. -
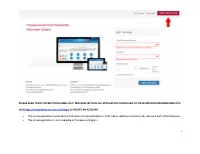
1 Please Read These Instructions Carefully
PLEASE READ THESE INSTRUCTIONS CAREFULLY. MISTAKES IN YOUR CSC APPLICATION COULD LEAD TO YOUR APPLICATION BEING REJECTED. Visit http://studyinchina.csc.edu.cn/#/login to CREATE AN ACCOUNT. • The online application works best with Firefox or Internet Explorer (11.0). Menu selection functions may not work with other browsers. • The online application is only available in Chinese and English. 1 • Please read this page carefully before clicking on the “Application online” tab to start your application. 2 • The Program Category is Type B. • The Agency No. matches the university you will be attending. See Appendix A for a list of the Chinese university agency numbers. • Use the + by each section to expand on that section of the form. 3 • Fill out your personal information accurately. o Make sure to have a valid passport at the time of your application. o Use the name and date of birth that are on your passport. Use the name on your passport for all correspondences with the CLIC office or Chinese institutions. o List Canadian as your Nationality, even if you have dual citizenship. Only Canadian citizens are eligible for CLIC support. o Enter the mailing address for where you want your admission documents to be sent under Permanent Address. Leave Current Address blank. Contact your home or host university coordinator to find out when you will receive your admission documents. Contact information for you home university CLIC liaison can be found here: http://clicstudyinchina.com/contact-us/ 4 • Fill out your Education and Employment History accurately. o For Highest Education enter your current degree studies.