Bulletin 110 FINAL EDIT.Indd
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Distribution and Habitat of the Invasive Giant Day Gecko Phelsuma Grandis Gray 1870
Phelsuma 22 (2014); 13-28 Distribution and habitat of the invasive giant day gecko Phelsuma grandis Gray 1870 (Sauria: Gekkonidae) in Reunion Island, and conservation implication Mickaël Sanchez1,* and Jean-Michel Probst1,2 1 Nature Océan Indien, 6, Lotissement les Magnolias, Rivière des Roches, 97470 Saint-Benoît, La Réunion, France 2 Nature et Patrimoine, 2, Allée Mangaron, Dos d’Ane, 97419 La Possession, La Réunion, France * Corresponding author; e-mail: [email protected] Abstract. The giant day gecko Phelsuma grandis, endemic to Madagascar, was introduced to Reunion Island (Indian Ocean) in the mid-1990s. No studies have been conducted so far to define its precise distribution and habitat. To fill the knowledge gap about this invasive species, we compiled available data and performed field work during 2007-2014. We detected 13 distinct populations of P. grandis, occurring mainly in the northern part of Reunion Island and on the west coast. This gecko inhabits human disturbed areas (gardens, urban parks, bamboos, orchards, coconuts, and banana plantation) and secondary habitats (shrubby savanna, and secondary dry woodlands, secondary dry and wet thickets). Its distribution strongly suggests that saltatory dispersal (through deliberate and/or accidental transport) and natural colonization are the mechanisms of spreading through Reunion Island. All our data in combination with both P. grandis ecology and native environmental range suggested that this gecko may colonize native forest, and constitutes a potential important threat to the native biodiversity of Reunion Island (arthropods and lizards). Key words. Phelsuma grandis, Reunion Island, distribution, invasive species, conservation. Introduction Day geckos of the genus Phelsuma are distributed in the western Indian Ocean (Austin et al. -

Market Snapshot Seychelles Hundred Islands of Paradise
FEBRUARY 2018 PRICE: US$250 MARKET SNAPSHOT SEYCHELLES HUNDRED ISLANDS OF PARADISE Larissa Lam Manager Daniel J Voellm Managing Partner HVS.com HVS | Level 7, Nan Fung Tower, 88 Connaught Road, Central, Hong Kong Seychelles Tourism Records Stable Growth Seychelles continues to prove its popularity in the European market as a hideaway holiday destination while gaining reputation in the Middle Eastern and Asian markets. Republic of Seychelles is an archipelago of 116 islands located in the Western Indian Ocean east of East Africa. Across all 116 islands, Seychelles has a total land area of 455 square kilometers. The country consists of two distinct island groups: the Mahé group, in the north, and a chain of low-lying coral islands stretching away to the south. All of the country’s principal islands belong to the Mahé group; they include Mahé Island (the largest at 27 kilometers long and 11 kilometers wide), Praslin Island, Silhouette, La Digue and 28 smaller islands. The 83 coral islands are largely without water resources, and most are uninhabited. The population of Seychelles (2017 estimate) is 93,920. Victoria, on Mahé Island, is the capital, principal city and leading port of Seychelles. While more than 75% of the country’s population lives on Mahé Island, about 10% on Praslin, and others on La Digue and the outer islands. ANSE INTENDANCE, MAHÉ Visitor Arrivals Seychelles is known for its coral beaches, opportunities for water sports, UNESCO heritage sites, its abundant wildlife and nature reserves and its year-long tropical climate. The government of Seychelles strictly controls tourism development in the country to protect and conserve the fragile ecosystem of the islands, and also to enable the long-term sustainability of the sector, which depends heavily on the country’s MARKET SNAPSHOT: SEYCHELLES - HUNDRED ISLANDS OF PARADISE | PAGE 2 ecological offerings. -

Secret Seychelles Islands with Ponant Aboard Le Jacques Cartier
SECRET SEYCHELLES ISLANDS WITH PONANT ABOARD LE JACQUES CARTIER Embark with PONANT on an expedition cruise to discover the most beautiful islands of the Seychelles. This 13-day itinerary aboard Le Jacques-Cartier will be an opportunity to discover little-known places of breathtaking natural beauty and an original fauna and flora. Leaving from Victoria, the archipelago’s capital, fall under the spell of the idyllic landscapes, with their exceptional flora and fauna. In Praslin, don’t miss the chance to visit the Vallée de Mai Nature Reserve. There you will find sea coconuts, gigantic fruits with a very evocative shape, nicknamed the “love nut”. You will discover the island of Aride, an unspoiled delight of the Indian Ocean, home to thousands of birds including some endemic species. During your cruise, you will have many opportunities to dive or snorkel, notably in Poivre, Assomption, Astove, and at the heart of the sublime coral reef in the Alphonse lagoon. Another highlight of your trip will be the port of call at Cosmoledo. This magnificent atoll owes its nickname, the Galapagos of the Indian Ocean, to the beauty of its unique underwater world. Diving in this paradise lagoon becomes an extraordinary experience. Before you return to Mahé, Le Jacques-Cartier will chart a course for the coral island of Desroches and the sublime beaches of La Digue, some of the most renowned of the Seychelles. The encounters with the wildlife described above illustrate possible experiences ITINERARY only and cannot be guaranteed. Day 1 VICTORIA, MAHÉ Discover Mahé, the main island of the Seychelles and also the largest of the archipelago, home to the capital, Victoria. -

Literature Cited in Lizards Natural History Database
Literature Cited in Lizards Natural History database Abdala, C. S., A. S. Quinteros, and R. E. Espinoza. 2008. Two new species of Liolaemus (Iguania: Liolaemidae) from the puna of northwestern Argentina. Herpetologica 64:458-471. Abdala, C. S., D. Baldo, R. A. Juárez, and R. E. Espinoza. 2016. The first parthenogenetic pleurodont Iguanian: a new all-female Liolaemus (Squamata: Liolaemidae) from western Argentina. Copeia 104:487-497. Abdala, C. S., J. C. Acosta, M. R. Cabrera, H. J. Villaviciencio, and J. Marinero. 2009. A new Andean Liolaemus of the L. montanus series (Squamata: Iguania: Liolaemidae) from western Argentina. South American Journal of Herpetology 4:91-102. Abdala, C. S., J. L. Acosta, J. C. Acosta, B. B. Alvarez, F. Arias, L. J. Avila, . S. M. Zalba. 2012. Categorización del estado de conservación de las lagartijas y anfisbenas de la República Argentina. Cuadernos de Herpetologia 26 (Suppl. 1):215-248. Abell, A. J. 1999. Male-female spacing patterns in the lizard, Sceloporus virgatus. Amphibia-Reptilia 20:185-194. Abts, M. L. 1987. Environment and variation in life history traits of the Chuckwalla, Sauromalus obesus. Ecological Monographs 57:215-232. Achaval, F., and A. Olmos. 2003. Anfibios y reptiles del Uruguay. Montevideo, Uruguay: Facultad de Ciencias. Achaval, F., and A. Olmos. 2007. Anfibio y reptiles del Uruguay, 3rd edn. Montevideo, Uruguay: Serie Fauna 1. Ackermann, T. 2006. Schreibers Glatkopfleguan Leiocephalus schreibersii. Munich, Germany: Natur und Tier. Ackley, J. W., P. J. Muelleman, R. E. Carter, R. W. Henderson, and R. Powell. 2009. A rapid assessment of herpetofaunal diversity in variously altered habitats on Dominica. -

Desroches-Island.Pdf
2012 FACT SHEET AMIRANTES GROUP OF ISLANDS I SEYCHELLES www.desroches-island.com JANUARY 2012 Location Desroches forms part of the Amirantes group of islands (a remote coral atoll), 230km south-west of Mahé in the Seychelles. Strewn with coconut trees, Desroches Island is approximately 6 km long, 1 km wide and boasts 14km of immaculate, white soft sandy beaches and crystal blue waters. This exclusive island is one of the most beautiful in the Indian Ocean. The island has only one luxurious resort with 20 Beach Suites and 26 Villas and 8 Retreats for guests to enjoy, all with sea-views and just a few steps from the beach. Located only 5˚ south of the equator Desroches is blessed with one of the world’s healthiest, malaria -free climates. Desroches is 10 hrs from London, 9 hrs from Paris, 8 hrs from Italy, 5 hrs from Johannesburg or Doha (Qatar) and 4 hrs from Dubai. BEACH SUITES Located around the central area, dining room, swimming pool and spa, the 20 luxuriously spacious BEACH SUITES BEACH Beach Suites are ideally built along the west-facing beach. Set amongst tropical gardens, palm trees and green lawns, the Beach Suites are just a few metres from the beach. – Each air-conditioned beach suite offers privacy with its own veranda, lounge area, flat screen TV and mini bar. Well designed bathrooms feature double basins, a covered outdoor bath, an outdoor shower and a separate toilet and bidet. To add to your exclusive experience, a choice of either in-suite dining or private dinners on the beach, where the waves lap gently against the soft white sand; are available. -

Atoll Research Bulletin No. 365 Issued by National
ATOLL RESEARCH BULLETIN NO. 365 ISSUED BY NATIONAL MUSEUM OF NATU HISTORY SMITHSONIAN INSTITUTION WASHINGTON, D.C., U.S.A. June 1992 CHAPTER 1 INTRODUCTION TO THE SOVIET- ERICAN EXPEDITION TO THE SEUCHELLES ISWDS The First Soviet-American Expedition in arine Biology to the Seychelles Islands was organized by the Institute of arine Biobgy, Far East Branc of the USSR Academy of Sciences, at the request of the SeycheIles Government. The Republic of the Seychelles Islands sought informari on the marine plane resources and productivity of benthic and planktonic corn Seychelles Bank. After discussing the program of w rrnent of Development of the Republic of the Seychelles Islands with the Deputy lvine Jandron, the following research objectives were established for the expedition: 1) To study the benthic marine biota and fouling processes of the Seychelles Islands, which represents a poorly investigated region of the Indian Ocean. 2) To provide the first analyses of the species composition of algal communities for several remote island groups (Farquhar Atoll, CBeeivy Atoll, Cosmoledo Atoll, hirantes Group) and to supplement previous knowledge on the algae of Aldabra Atoll, ah6 Island and Praslin Island. 3) To study the distribution of autotrophic organisms over the various reef systems and determine the depth ranges of algae, seagrasses and corals. 4) To evaluate the common algal and seagrass resources in the area of study, particularly species of commercial interest. 5) To estimate the production potential of the major producers of organic matter on Seychelles reefs; i.e., benthic macroalgae, seagrasses, reef building corals and phytoplankton. 6) To assess the prevalent environmental parameters of the various island groups studied: e.g., optical characteristics of the water, seawater temperatures, nutrient contents, oxygen levels, pH and current velocities. -

1 7 an Identification Key to the Geckos of the Seychelles
HERPETOLOGICAL JOURNAL. Vol. I. pp. 17-19 (19X5l 17 AN IDENTIFICATION KEY TO THE GECKOS OF THE SEYCHELLES, WITH BRIEF NOTES ON THEIR DISTRIBUTIONS AND HABITS ANDREW S. GARDNER Department of Zoology, University of Aberdren. Ti/lydrone Avenue, Aberdeen AB9 2TN. U. K. Present addresses: The Calton Laboratory. Department of Genetics and Biomet IT, Universif.I' Co/legr London. Wo/f�·on !-louse, 4 Stephenson Wa r London NWI 21-11'.. U.K. (A ccepted 24. /0. 84) INTRODUCTION 4. Scales on chest and at least anterior of belly keeled. Underside white. Phe!suma astriata The Republic of Seychelles, lying in the western Tornier. 5. Indian Ocean consists of a group of mountainous, granitic islands, and a large number of outlying coral Scales on chest and belly not keeled. 6. atolls and sand cays, distributed over 400,000 km2 of sea. There are over a hundred islands, ranging in size 5. Subcaudal scales keeled and not transversely from Mahe, at 148 km2 to islands little more than enlarged in original tails. Ground colour of emergent rocks. A total of eighteen species of lizard, rump and tail usually bright blue, and of from three families are recorded from the Seychelles nanks, green. Tail unmarked or spotted with (Gardner, 1984). The best represented family is the red. Red transverse neck bars often reduced or Gekkonidae with eleven species, fo ur of which are absent. Phe/suma astriata astriata Tornier i endemic to the islands. The identification key 90 1. presented here should enable interested naturalists to Subcaudal scales unkeeled and transversely identify any gecko encountered in the Seychelles to the enlarged in original tails. -

Atoll Research Bulletin No. 252 Bird and Denis Islands
ATOLL RESEARCH BULLETIN NO. 252 BIRD AND DENIS ISLANDS, SEYCHELLES by D. R. Stoddart and F. R. Fosberg Issued by THE SMITHSONIAN INSTITUTION Washington, D. C., U.S.A. ~ul~'l981 Contents 1. Geography and ecology of Bird Island, Seychelles Introduction Morphology and structure Climate Vegetation Flora Invertebrates Reptiles Mammals Birds History 2. Plants recorded from Bird Island 3. Geography and ecology of Denis Island, Seychelles Introduction Morphology and structure Climate Vegetation Flora Invertebrates Reptiles Mammals Birds History 4. Plants recorded from Denis Island 5. References Manuscript received May 1980 --Eds. List of Figures 1. The Seychelles Bank following page 11 2. Bird Island in 1976 following page 11 3. Beach sediment at Bird Island following page 11 4. Denis Island in 1977 following page 50 5. Monthly rainfall at Denis Island, 19 71-1962 following page 50 List of Tables 1. Scientific studies at Bird Island 2. Characteristics of Bird Island beach sands 3. Monthly rainfall at Bird Island, 1951-1962 4. Key to the literature on insects collected at Bird Island 5. Scientific studies at Denis Island 6. Monthly and annual rainfall records at Denis Island iii List of Plates Bird Island: Suriana zone on the northeast shore following page 11 Bird Island: Pisonia and Cordia woodland with Suriana on the northeast shore Bird Island: Tournefortia parkland in the northeast Bird Island: tree-like Tournefortia in the northeast Bird Island: pioneer sedges and Scaevola on the east shore Bird Island: pioneer Ipomoea pes-caprae on the east shore Bird Island: pioneer sedges, Scaevola and Tournefortia on the northeast shore Bird Island: airstrip from the southeast Denis Island: phosphate cliffs with Casuarina woodland, southwest shore following page 50 10. -

Aldabra Atoll the Seychelles Is an Archipelago in the Western Indian Ocean, Spread out Within an Exclusive Economic Zone (EEZ) of 1.3 Million Km2
CONTENTS INTRODUCTION.............................................................................................................................................1 HOW THE EVALUATION WAS CARRIED OUT..........................................................................................4 CONTEXT REVIEW Focal Management Targets .........................................................................................................................6 • Data sheet ...............................................................................................................................................8 Threats to World Heritage Values and Focal Management Targets....................................................11 • Data sheet .............................................................................................................................................13 Review of National Context .......................................................................................................................16 • Data sheet .............................................................................................................................................18 Engagement of stakeholders and partners.............................................................................................23 • Data sheets ...........................................................................................................................................26 PLANNING ASSESSMENT Management Planning Assessment.........................................................................................................38 -

Roxtons-Blue-Safari-Brochure.Pdf
25 High Street, Hungerford, Berkshire RG17 0NF England Tel: +44 (0)1488683222 Email: [email protected] Website: www.roxtons.com THE SHEER REMOTENESS OF THE OUTER ISLANDS ARE Mahé ABUNDANT WITH AN EXTRAORDINARY SELECTION OF WILDLIFE, FLORA AND FAUNA, CULMINATING IN AN 40 min UNPARALLELED BIOME OF DIVERSITY. Poivre Atoll 60 min Desroches Island These magnificent islands and atolls, consisting of the Alphonse Group of Islands – the stunning trio of Alphonse Island, St François Island and Bijoutier Island and the other beautiful atolls of Cosmoledo Atoll, Astove 30 min Atoll, Poivre Atoll and the resplendent Amirante Islands are found south Cosmoledo Atoll of the equator in the heart of the Indian Ocean. These range between 400 and 1 055 kilometres south-west of Mahé, the main island of the Seychelles. Alphonse Island They are blessed with one of the world’s healthiest climates, with miles of unblemished shorelines, lagoons and sea flats. The isolation of these islands and atolls creates an atmosphere of undisturbed solitude where you Bijoutier Island can immerse yourself in the natural wonders and exceptional experiences 60 min they have to ofer. 120 min Considered some of the untouched Edens of the world, these outer islands and atolls of the Seychelles have an unparalleled biome of diversity and St François Island Astove Atoll abundance, making them a nature lover’s paradise which begs to be dis- covered and explored. What is Blue Safari Seychelles? Safari is a Swahili word taken from the Arabic, safar ‘to journey’. Blue is the predominant colour in the area, and if it’s your time to journey into the unknown; to seek out excitement and adventure, to walk amongst mangroves, free-living giant land tortoises and wild spaces, then read on.. -
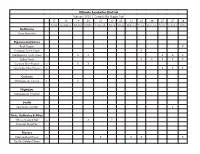
Ultimate Seychelles Bird List February 2020 | Compiled by Pepper Trail
Ultimate Seychelles Bird List February 2020 | Compiled by Pepper Trail 17 18 19 20 21 22 23 24 25 26 27 27 28 At Sea Assumption Aldabra Aldabra Cosmoledo Astove At Sea Alphonse Poivre Desroches Praslin La Digue Mahe Galliforms Gray Francolin Pigeons and Doves Rock Pigeon X European Turtle-Dove X Madagascar Turtle-Dove x X X X X X Zebra Dove X X X X X Comoro Blue-Pigeon x X X Seychelles Blue-Pigeon x X X X Cuckoos Madagascar Coucal x X Nightjars Madagascar Nightjar Swifts Seychelles Swiftlet x X Rails, Gallinules & Allies White-throated Rail x X ? Eurasian Moorhen Plovers Black-bellied Plover x X X X X Pacific Golden-Plover Ultimate Seychelles Bird List February 2020 | Compiled by Pepper Trail 17 18 19 20 21 22 23 24 25 26 27 27 28 At Sea Assumption Aldabra Aldabra Cosmoledo Astove At Sea Alphonse Poivre Desroches Praslin La Digue Mahe Lesser Sand-Plover x X Greater Sand-Plover x X X Common Ringed Plover Sandpipers Whimbrel x X X X X X X Eurasian Curlew Bar-tailed Godwit Ruddy Turnstone x X X X X X Curlew Sandpiper x X X Sanderling x X X Little Stint Terek Sandpiper Common Sandpiper x X Common Greenshank x X X X Wood Sandpiper Crab-Plover x X X X X X Gulls and Terns Brown Noddy x X X X X X X X X Lesser Noddy x X White Tern x X X X X X X X X Sooty Tern Bridled Tern x X X X Saunders’s Tern White-winged Tern Roseate Tern Ultimate Seychelles Bird List February 2020 | Compiled by Pepper Trail 17 18 19 20 21 22 23 24 25 26 27 27 28 At Sea Assumption Aldabra Aldabra Cosmoledo Astove At Sea Alphonse Poivre Desroches Praslin La Digue Mahe Black-naped -

PRE-TRIP INFORMATION Astove Atoll WELCOME to YOUR ULTIMATE FISHING EXPERIENCE
PRE-TRIP INFORMATION Astove Atoll WELCOME TO YOUR ULTIMATE FISHING EXPERIENCE... www.alphonsefishingco.com JANUARY 2020 Frankfurt Dusseldorf London INTERNATIONAL FLIGHTS Paris Vienna Zurich Istanbul DIRECT FLIGHTS TO/FROM SEYCHELLES Abu Dhabi Doha Dubai Bombay EQUATOR Addis Ababa Colombo Nairobi Seychelles Dar-Es-Salaam Mauritius Johannesburg Reunion MAHÉ 40 min St Joseph Island D’Arros Island Desroches Island Poivre Island Astove Atoll 60 min 30 min Astove is situated 1055 km southwest of Mahé and forms part of the remote Aldabra group of atolls. It’s a small and unique atoll that spans six kilometers from north to south and just under four kilometers Alphonse Island from east to west at the widest points. The shallow lagoon has one small entrance, and due to its elevation a phenomenon occurs whereby the tide falls like a river for ten hours of the 12-hour tidal cycle and then turns to flood the entire lagoon in only two hours. 120 min ASTOVE CORAL HOUSE Aldabra Atoll Cosmoledo Atoll Assumption Island ASTOVE ISLAND ASTOVE ATOLL LAGOON www.alphonsefishingco.com 1 ASTOVE ATOLL The Wildest Fly Fishing Destination on this Planet! WHAT YOU CAN EXPECT FISHING AT ASTOVE ATOLL Astove has become synonymous with the largest flats-caught GT’s in Not only does Astove have large GT’s but it also has an equally the Indian Ocean. Its shallow lagoon and small entrance, surrounded impressive bonefish, permit, bluefin trevally, triggerfish, barracuda by sheer drop-offs makes the experience truly unique. and milkfish population. This lagoon is a sanctuary for both juvenile and trophy-size fish that The offshore fishing starts metres from the edge of the flats where feed on the shallow white sand flats that line the inside of the lagoon.