Vegetation Units Within the West Angelas Study Area Map D2
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Additional Information
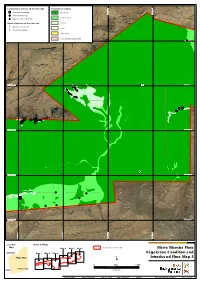
Current Survey Introduced Flora Records Vegetation Condition *Acetosa vesicaria Excellent 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 -

Seed Ecology Iii
SEED ECOLOGY III The Third International Society for Seed Science Meeting on Seeds and the Environment “Seeds and Change” Conference Proceedings June 20 to June 24, 2010 Salt Lake City, Utah, USA Editors: R. Pendleton, S. Meyer, B. Schultz Proceedings of the Seed Ecology III Conference Preface Extended abstracts included in this proceedings will be made available online. Enquiries and requests for hardcopies of this volume should be sent to: Dr. Rosemary Pendleton USFS Rocky Mountain Research Station Albuquerque Forestry Sciences Laboratory 333 Broadway SE Suite 115 Albuquerque, New Mexico, USA 87102-3497 The extended abstracts in this proceedings were edited for clarity. Seed Ecology III logo designed by Bitsy Schultz. i June 2010, Salt Lake City, Utah Proceedings of the Seed Ecology III Conference Table of Contents Germination Ecology of Dry Sandy Grassland Species along a pH-Gradient Simulated by Different Aluminium Concentrations.....................................................................................................................1 M Abedi, M Bartelheimer, Ralph Krall and Peter Poschlod Induction and Release of Secondary Dormancy under Field Conditions in Bromus tectorum.......................2 PS Allen, SE Meyer, and K Foote Seedling Production for Purposes of Biodiversity Restoration in the Brazilian Cerrado Region Can Be Greatly Enhanced by Seed Pretreatments Derived from Seed Technology......................................................4 S Anese, GCM Soares, ACB Matos, DAB Pinto, EAA da Silva, and HWM Hilhorst -

The-Potential-Use-For-Groundwater
i Professor Peter Cook 84 Richmond Avenue Colonel Light Gardens SA 5041 [email protected] Professor Derek Eamus School of Life Sciences University of Technology Sydney PO Box 123 Sydney NSW 2007 [email protected] Cover Photo: Open woodland vegetation in the Ti Tree Basin. ii Table of Contents Executive Summary .................................................................................................................... v 1. INTRODUCTION ................................................................................................................... 9 2. METHODOLOGIES TO INFER GROUNDWATER USE .......................................................... 11 2.1 Direct Measurements of Rooting Depth 11 2.2 Soil Water Potentials 12 2.3 Leaf and Soil Water Potentials 13 2.4 Stable Isotopes 2H and 18O 14 2.5 Depth of Water Use and Groundwater Access 16 2.6 Green Islands 17 2.7 Transpiration Rates 19 2.8 Tree Rings 20 2.9 Dendrometry 22 2.10 13C of Sapwood 22 3. GROUNDWATER AND VEGETATION IN THE TI TREE BASIN .............................................. 24 3.1 Geography and Climate 24 3.2 Groundwater Resources 27 3.3 Vegetation Across the Ti Tree Basin 29 4. TI TREE BASIN GDE STUDIES ............................................................................................. 32 4.1 Transpiration and Evapotranspiration Rates 32 4.2 Soil Water Potentials 35 4.3 Leaf Water Potentials 38 4.4 Stable Isotopes 43 4.5 Sapwood 13C and Leaf Vein Density 44 5. OTHER ARID ZONE STUDIES ............................................................................................. -

Handbook of Grasses, Treating of Their Structure, Classification
Young collector ."9' 910 JAN: W„HUTCHINSON ONE SEIinNG BOUGHT WITH THE INCOME OF THE SAGE ENDOWMENT FUND THE GIFT OF HENRY W. SAGE 1891 ALBERT R. MANN LIBRARY New York State Colleges OF Agriculture and Human Ecology AT Cornell University Cornell University Library QK 495.G74H97 1910 •'e''*'"9 Handbook of grasses, "UJimirn^' 3 1924 001 738 933 Cornell University Library The original of tliis book is in tlie Cornell University Library. There are no known copyright restrictions in the United States on the use of the text. http://www.archive.org/details/cu31924001738933 Poa annua, the Annual Meadow-grass ; flowering. This Book is now publislitd by GEORGE ALLEN' & UNWIN, LTD. Ruskiii House, 40, MUSEUM STREET, LONDON^ W.C. Poa anmia, the Annual Meadow-grassj floweidns HANDBOOK OF GRASSES TREATING OF THEIR STRUCTURE, CLASSIFICATION, GEOGRAPHICAL DISTRIBUTION AND USES ALSO DESCRIBING THE BRITISH SPECIES AND THEIR HABITATS BY WILLIAM HUTCHINSON XonOon SONNENSCHEIN & CO., Lim. , SWAN New York : THE MACMILLAN CO. igio Edition, March iSgg; Third Edition igob; First Edition, SeftembeiiSgs; Second Fourth Edition, igio. PREFACE Grasses are in three respects a remarkable family : they possess many structural peculiarities which sharply define them from all other kinds of plants ; they are so abundant and widely diffused as to constitute the dominant feature of the landscape, not only in our own, but in most other coun- tries j and lastly, no other Order can at all compare with the Gramineffi in the variety and magnitude of their uses. Yet the study of grasses, so far from being popular, is shunned by many botanists in the belief that it is beset with unusual difficulties ; farmers and graziers, to whom the cereal and forage grasses are all in all, have rarely a scientific acquaintance with them ; while those observers of Nature, not particularly iriterested in either botany or agriculture, are hardly able to recognize two or three among the many species which everywhere abound. -

Climate Teleconnections Synchronize Picea Glauca Masting and Fire Disturbance: Evidence for a Fire‐Related Form of Environmental Prediction
Received: 9 August 2019 | Accepted: 7 October 2019 DOI: 10.1111/1365-2745.13308 RESEARCH ARTICLE Climate teleconnections synchronize Picea glauca masting and fire disturbance: Evidence for a fire‐related form of environmental prediction Davide Ascoli1 | Andrew Hacket‐Pain2 | Jalene M. LaMontagne3 | Adrián Cardil4 | Marco Conedera5 | Janet Maringer5 | Renzo Motta1 | Ian S. Pearse6 | Giorgio Vacchiano7 1Department of Agricultural, Forestry and Food Sciences, University of Torino, Grugliasco, Italy; 2Department of Geography and Planning, School of Environmental Sciences, University of Liverpool, Liverpool, UK; 3Department of Biological Sciences, DePaul University, Chicago, IL, USA; 4Department of Crops and Forest Sciences, University of Lleida, Lleida, Spain; 5Insubric Ecosystems, Swiss Federal Institute for Forest, Snow and Landscape Research WSL, Cadenazzo, Switzerland; 6Fort Collins Science Center, U.S. Geological Survey, Fort Collins, CO, USA and 7Department of Agricultural and Environmental Sciences, University of Milan, Milan, Italy Correspondence Davide Ascoli Abstract Email: [email protected] 1. Synchronous pulses of seed masting and natural disturbance have positive feed- Funding information backs on the reproduction of masting species in disturbance-prone ecosystems. Natural Environment Research Council, We test the hypotheses that disturbances and proximate causes of masting are Grant/Award Number: NE/S007857/1; NSF, Grant/Award Number: DEB-1745496 correlated, and that their large-scale synchrony is driven by similar climate tel- econnection patterns at both inter-annual and decadal time scales. Handling Editor: Peter Bellingham 2. Hypotheses were tested on white spruce (Picea glauca), a masting species which surprisingly persists in fire-prone boreal forests while lacking clear fire adap- tations. We built masting, drought and fire indices at regional (Alaska, Yukon, Alberta, Quebec) and sub-continental scales (western North America) spanning the second half of the 20th century. -

Kingdom Class Family Scientific Name Common Name I Q a Records
Kingdom Class Family Scientific Name Common Name I Q A Records plants monocots Poaceae Paspalidium rarum C 2/2 plants monocots Poaceae Aristida latifolia feathertop wiregrass C 3/3 plants monocots Poaceae Aristida lazaridis C 1/1 plants monocots Poaceae Astrebla pectinata barley mitchell grass C 1/1 plants monocots Poaceae Cenchrus setigerus Y 1/1 plants monocots Poaceae Echinochloa colona awnless barnyard grass Y 2/2 plants monocots Poaceae Aristida polyclados C 1/1 plants monocots Poaceae Cymbopogon ambiguus lemon grass C 1/1 plants monocots Poaceae Digitaria ctenantha C 1/1 plants monocots Poaceae Enteropogon ramosus C 1/1 plants monocots Poaceae Enneapogon avenaceus C 1/1 plants monocots Poaceae Eragrostis tenellula delicate lovegrass C 2/2 plants monocots Poaceae Urochloa praetervisa C 1/1 plants monocots Poaceae Heteropogon contortus black speargrass C 1/1 plants monocots Poaceae Iseilema membranaceum small flinders grass C 1/1 plants monocots Poaceae Bothriochloa ewartiana desert bluegrass C 2/2 plants monocots Poaceae Brachyachne convergens common native couch C 2/2 plants monocots Poaceae Enneapogon lindleyanus C 3/3 plants monocots Poaceae Enneapogon polyphyllus leafy nineawn C 1/1 plants monocots Poaceae Sporobolus actinocladus katoora grass C 1/1 plants monocots Poaceae Cenchrus pennisetiformis Y 1/1 plants monocots Poaceae Sporobolus australasicus C 1/1 plants monocots Poaceae Eriachne pulchella subsp. dominii C 1/1 plants monocots Poaceae Dichanthium sericeum subsp. humilius C 1/1 plants monocots Poaceae Digitaria divaricatissima var. divaricatissima C 1/1 plants monocots Poaceae Eriachne mucronata forma (Alpha C.E.Hubbard 7882) C 1/1 plants monocots Poaceae Sehima nervosum C 1/1 plants monocots Poaceae Eulalia aurea silky browntop C 2/2 plants monocots Poaceae Chloris virgata feathertop rhodes grass Y 1/1 CODES I - Y indicates that the taxon is introduced to Queensland and has naturalised. -

DRAFT 25/10/90; Plant List Updated Oct. 1992; Notes Added June 2021
DRAFT 25/10/90; plant list updated Oct. 1992; notes added June 2021. PRELIMINARY REPORT ON THE CONSERVATION VALUES OF OPEN COUNTRY PADDOCK, BOOLARDY STATION Allan H. Burbidge and J.K. Rolfe INTRODUCTION Boolardy Station is situated about 150 km north of Yalgoo and 140 km west-north-west of Cue, in the Shire of Murchison, Western Australia. Open Country Paddock (about 16 000 ha) is in the south-east corner of the station, at 27o05'S, 116o50'E. The most prominent named feature is Coolamooka Hill, near the eastern boundary of the paddock. There are no conservation reserves in this region, although there are some small reserves set aside for various other purposes. Previous biological data for the station consist of broad scale vegetation mapping and land system mapping. Beard (1976) mapped the entire Murchison region at 1: 1 000 000. The Open Country Paddock area was mapped as supporting mulga woodlands and shrublands. More detailed mapping of land system units for rangeland assessment purposes has been carried out more recently at a scale of 1: 40 000 (Payne and Curry in prep.). Seven land systems were identified in open Country Paddock (Fig. 1). Apart from these studies, no detailed biological survey work appears to have been done in the area. Open Country Paddock has been only lightly grazed by domestic stock because of the presence of Kite-leaf Poison (Gastrolobium laytonii) and a lack of fresh water. Because of this and the generally good condition of the paddock and presence of a wide range of plant species, P.J. -

Enabling the Market: Incentives for Biodiversity in the Rangelands
Enabling the Market: Incentives for Biodiversity in the Rangelands: Report to the Australian Government Department of the Environment and Water Resources by the Desert Knowledge Cooperative Research Centre Anita Smyth Anthea Coggan Famiza Yunus Russell Gorddard Stuart Whitten Jocelyn Davies Nic Gambold Jo Maloney Rodney Edwards Rob Brandle Mike Fleming John Read June 2007 Copyright and Disclaimers © Commonwealth of Australia 2007 Information contained in this publication may be copied or reproduced for study, research, information or educational purposes, subject to inclusion of an acknowledgment of the source. The views and opinions expressed in this publication are those of the authors and do not necessarily reflect those of the Australian Government or the Minister for the Environment and Water Resources. While reasonable efforts have been made to ensure that the contents of this publication are factually correct, the Australian Government does not accept responsibility for the accuracy or completeness of the contents, and shall not be liable for any loss or damage that may be occasioned directly or indirectly through the use of, or reliance on, the contents of this publication. Contributing author information Anita Smyth: CSIRO Sustainable Ecosystems Anthea Coggan: CSIRO Sustainable Ecosystems Famiza Yunus: CSIRO Sustainable Ecosystems Russell Gorddard: CSIRO Sustainable Ecosystems Stuart Whitten: CSIRO Sustainable Ecosystems Jocelyn Davies: CSIRO Sustainable Ecosystems Nic Gambold: Central Land Council Jo Maloney Rodney Edwards: Ngaanyatjarra Council Rob Brandle: South Austalia Department for Environment and Heritage Mike Fleming: South Australia Department of Water, Land and Biodiversity Conservation John Read: BHP Billiton Desert Knowledge CRC Report Number 18 Information contained in this publication may be copied or reproduced for study, research, information or educational purposes, subject to inclusion of an acknowledgement of the source. -

By H.D.V. PRENDERGAST a Thesis Submitted for the Degree of Doctor of Philosophy of the Australian National University. January 1
STRUCTURAL, BIOCHEMICAL AND GEOGRAPHICAL RELATIONSHIPS IN AUSTRALIAN c4 GRASSES (POACEAE) • by H.D.V. PRENDERGAST A thesis submitted for the degree of Doctor of Philosophy of the Australian National University. January 1987. Canberra, Australia. i STATEMENT This thesis describes my own work which included collaboration with Dr N .. E. Stone (Taxonomy Unit, R .. S .. B.S .. ), whose expertise in enzyme assays enabled me to obtain comparative information on enzyme activities reported in Chapters 3, 5 and 7; and with Mr M.. Lazarides (Australian National Herbarium, c .. s .. r .. R .. O .. ), whose as yet unpublished taxonomic views on Eragrostis form the basis of some of the discussion in Chapter 3. ii This thesis describes the results of research work carried out in the Taxonomy Unit, Research School of Biological Sciences, The Australian National University during the tenure of an A.N.U. Postgraduate Scholarship. iii ACKNOWLEDGEMENTS My time in the Taxonomy Unit has been a happy one: I could not have asked for better supervision for my project or for a more congenial atmosphere in which to work. To Dr. Paul Hattersley, for his help, advice, encouragement and friendship, I owe a lot more than can be said in just a few words: but, Paul, thanks very much! To Mr. Les Watson I owe as much for his own support and guidance, and for many discussions on things often psittacaceous as well as graminaceous! Dr. Nancy Stone was a kind teacher in many days of enzyme assays and Chris Frylink a great help and friend both in and out of the lab •• Further thanks go to Mike Lazarides (Australian National Herbarium, c.s.I.R.O.) for identifying many grass specimens and for unpublished data on infrageneric groups in Eragrostis; Dr. -

Northwest Highlands Bioregion Technical Descriptions
Department of Environment and Science Regional Ecosystem Technical Descriptions Technical descriptions provide a detailed description of the full range in structure and floristic composition of regional ecosystems (e.g. 1.1.10) and their component vegetation communities (e.g. 1.11.10a, 1.11.10b). The descriptions are compiled using site survey data from the Queensland Herbarium’s CORVEG database. Distribution maps, representative images (if available) and the pre-clearing and remnant area (hectares) of each vegetation community derived from the regional ecosystem mapping (spatial) data are included. The technical descriptions should be used in conjunction with the fields from the regional ecosystem description database (REDD) for a full description of the regional ecosystem. Quantitative site data from relatively undisturbed sites are extracted from CORVEG and summarized to provide information specific to each vegetation community. Technical descriptions include the attributes: tree canopy height and cover and native plant species composition of the predominant layer, which are used to assess the remnant status of vegetation under the Vegetation Management Act 1999. However, as technical descriptions reflect the full range in structure and floristic composition across the climatic, natural disturbance and geographic range of the regional ecosystem, local reference sites should be used where possible (Neldner et al. 2005 section 3.3.3). The technical descriptions are subject to review and are updated as additional data becomes available. -

Flora Survey on Hiltaba Station and Gawler Ranges National Park
Flora Survey on Hiltaba Station and Gawler Ranges National Park Hiltaba Pastoral Lease and Gawler Ranges National Park, South Australia Survey conducted: 12 to 22 Nov 2012 Report submitted: 22 May 2013 P.J. Lang, J. Kellermann, G.H. Bell & H.B. Cross with contributions from C.J. Brodie, H.P. Vonow & M. Waycott SA Department of Environment, Water and Natural Resources Vascular plants, macrofungi, lichens, and bryophytes Bush Blitz – Flora Survey on Hiltaba Station and Gawler Ranges NP, November 2012 Report submitted to Bush Blitz, Australian Biological Resources Study: 22 May 2013. Published online on http://data.environment.sa.gov.au/: 25 Nov. 2016. ISBN 978-1-922027-49-8 (pdf) © Department of Environment, Water and Natural Resouces, South Australia, 2013. With the exception of the Piping Shrike emblem, images, and other material or devices protected by a trademark and subject to review by the Government of South Australia at all times, this report is licensed under the Creative Commons Attribution 4.0 International License. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/. All other rights are reserved. This report should be cited as: Lang, P.J.1, Kellermann, J.1, 2, Bell, G.H.1 & Cross, H.B.1, 2, 3 (2013). Flora survey on Hiltaba Station and Gawler Ranges National Park: vascular plants, macrofungi, lichens, and bryophytes. Report for Bush Blitz, Australian Biological Resources Study, Canberra. (Department of Environment, Water and Natural Resources, South Australia: Adelaide). Authors’ addresses: 1State Herbarium of South Australia, Department of Environment, Water and Natural Resources (DEWNR), GPO Box 1047, Adelaide, SA 5001, Australia. -

Rare Or Threatened Vascular Plant Species of Wollemi National Park, Central Eastern New South Wales
Rare or threatened vascular plant species of Wollemi National Park, central eastern New South Wales. Stephen A.J. Bell Eastcoast Flora Survey PO Box 216 Kotara Fair, NSW 2289, AUSTRALIA Abstract: Wollemi National Park (c. 32o 20’– 33o 30’S, 150o– 151oE), approximately 100 km north-west of Sydney, conserves over 500 000 ha of the Triassic sandstone environments of the Central Coast and Tablelands of New South Wales, and occupies approximately 25% of the Sydney Basin biogeographical region. 94 taxa of conservation signiicance have been recorded and Wollemi is recognised as an important reservoir of rare and uncommon plant taxa, conserving more than 20% of all listed threatened species for the Central Coast, Central Tablelands and Central Western Slopes botanical divisions. For a land area occupying only 0.05% of these divisions, Wollemi is of paramount importance in regional conservation. Surveys within Wollemi National Park over the last decade have recorded several new populations of signiicant vascular plant species, including some sizeable range extensions. This paper summarises the current status of all rare or threatened taxa, describes habitat and associated species for many of these and proposes IUCN (2001) codes for all, as well as suggesting revisions to current conservation risk codes for some species. For Wollemi National Park 37 species are currently listed as Endangered (15 species) or Vulnerable (22 species) under the New South Wales Threatened Species Conservation Act 1995. An additional 50 species are currently listed as nationally rare under the Briggs and Leigh (1996) classiication, or have been suggested as such by various workers. Seven species are awaiting further taxonomic investigation, including Eucalyptus sp.