Analyzing Mitochondria at Higher Resolution
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Hyperbranched Polyaspartate Esters and a Process for Their Preparation
Europäisches Patentamt *EP000743335B1* (19) European Patent Office Office européen des brevets (11) EP 0 743 335 B1 (12) EUROPEAN PATENT SPECIFICATION (45) Date of publication and mention (51) Int Cl.7: C08G 63/685 of the grant of the patent: 26.09.2001 Bulletin 2001/39 (21) Application number: 96107069.5 (22) Date of filing: 06.05.1996 (54) Hyperbranched polyaspartate esters and a process for their preparation Hyperverzweigte Polyaspartatester und Verfahren zu ihrer Herstellung Ester de polyaspartate hyperramifiés et procédé pour leur préparation (84) Designated Contracting States: (74) Representative: Pettrich, Klaus-Günter, Dr. AT BE CH DE ES FR GB IT LI NL SE c/o Bayer AG, Konzernbereich RP (30) Priority: 18.05.1995 US 443505 Patente und Lizenzen 51368 Leverkusen (DE) (43) Date of publication of application: 20.11.1996 Bulletin 1996/47 (56) References cited: US-A- 5 126 170 (73) Proprietor: Bayer Corporation Pittsburgh, PA 15205-9741 (US) • ISRA L JOURNAL OF CHEMISTRY, vol. 9, 1971, JERUSALEM, pages 105-109, XP000670606 A. (72) Inventors: SINGERMAN ET AL.: "Poly threo-beta-hydroxy • Yeske, Philip E. aspartic acid" Pittsburgh, PA 15228 (US) • Gindin, Lyuba K. Remarks: Pittsburgh, PA 15216 (US) The file contains technical information submitted • Wicks, Douglas A. after the application was filed and not included in this Mt. Lebanon, PA 15228 (US) specification • Jonsson, E. Haakan Coraopolis, PA 15108 (US) Note: Within nine months from the publication of the mention of the grant of the European patent, any person may give notice to the European Patent Office of opposition to the European patent granted. Notice of opposition shall be filed in a written reasoned statement. -

The Mechanism of Aconitase Action Iii. Kinetic Analysis Using Dl-Isocitric Acid-2-C 14
TheJournal of Biochemistry, Vol.41, No. 5, 1954 THE MECHANISM OF ACONITASE ACTION III. KINETICANALYSIS USING DL-ISOCITRIC ACID-2-C14 By JUN-ICHI TOMIZAWA (Fromthe NationalInstitute of Health,Tokyo) (Receivedfor publication,June 24, 1954). In the previous reports (1, 2, 3), the author came to a conclusion that aconitase would be a single enzyme, and one enzyme and one activated complex theory was proposed. In the present communication, some additional proofs of the theory using labeled substrates will be reported. EXPERIMENTAL The Enzyme Preparation and the Methods of Analysis The preparation of the rabbit liver enzyme was the same as reported previously (1) except it was centrifuged at 120,000 x g for 30 minutes and the supernatant was kept. General properties of the enzyme preparation were not changed by this treat ment. The preparation and the analysis of non-labeled substrates were previously reported. The Preparation of DL-Isocitric Acid-2-C14 Usually DL-isocitric acid was prepared by hydrolysis of trichloromethylparaconic acid. It was, however, rather difficult to prepare the carbon-2 labeled compound by this method. Therefore, the compound was prepared by an entirely new process. Ethyl Formate-C14•\Formic acid was prepared by the usual method from labeled barium carbonate. Its ethyl ester was prepared by esterification of the sodium salt as was done in the preparation of its methyl ester according to Me1viIIeetal. (4, 5). The yield was about 80per cent. Diethyl Formyl-succinate-l-C14-This compound was prepared by the method of Sugazawa (6). 0.6g. of ethyl formate-C14 and 1.2g. -
EFFECT of INOCULUM on KINETICS and YIELD of CITRIC ACIDS PRODUCTION on GLUCOSE by Yarrowia Lipolytica A-101
.\ C T A ALIMENTARIA POLONI C A Vol. XVII/ XLI ! No. 2 1991 MARIA WOJTATOWICZ WALDEMAR RYMOWICZ EFFECT OF INOCULUM ON KINETICS AND YIELD OF CITRIC ACIDS PRODUCTION ON GLUCOSE BY Yarrowia Lipolytica A-101 Departmcnt of Biotechnology and Food Microbiology, Academy of Agriculture, Wrocław Key word s: Yarrowia fipo(1 'fin,1 /\-1 O1 , citric and isodtric acid, glucosc, nitrogen deficient medium The cffcct of two differcnt inocula on growth and production parameters in citric acid fcrmcntation (on glucose) by Yarrowia lipolytica A-101 was studied. For inoculum prcpared in full growth medium the total acids yield was 5-12% higher and · biomass yield about 10 % higher than for inoculum prepared in a nitrogen-deficient medium. The latter inoeulum, however, !cd to about 10-30% highcr acid producti.on and glucosc consumption rates. Until the early I 970s practically the only organisms used to produce citric acid were Aspergillus niger and a few other fungi. Today we know that many kinds of yeasts can accumulate substantial amounts of citric acid in their growth media. The most cfficient citric acid producers belong to the Candida genus, and the strains used most often are C. lipolytica, C. zeylanoides, C. parapsilosis, C. tropicalis, C. guilliermondii, C. oleophila, C. petrophilum and C. intermedia [10]. Yeasts can produce citric acid more rapidly than fungi and in a greater variety of substrates including n-alkanes, n-alkenes, glucose, molasses, acetate, alcohols, fatty acids and natura) oils [4, 8, 9, 17, 18]. The product yield on n-alkanes and vegetable oils may be as high as 1.6 g/g [4, 9]; on glucose it is usually comparable to that in proccsses involving filamentous molds [4-6]. -

D-Isocitric Acid (D-Isocitrate)
www.megazyme.com D-ISOCITRIC ACID (D-ISOCITRATE) ASSAY PROCEDURE K-ISOC 11/19 (*100 Manual Assays per Kit) or (1000 Auto-Analyser Assays per Kit) or (1000 Microplate Assays per Kit) * The number of tests per kit can be doubled if all volumes are halved © Megazyme 2019 INTRODUCTION: D-Isocitric acid is an organic acid found in most fruit juices. It is an important marker in multicomponent procedures for the evaluation of authenticity and quality of fruit products; high citric/isocitric acid ratios can be used as an indicator of citric acid addition in some juices. PRINCIPLE: D-Isocitric acid is oxidised by nicotinamide-adenine dinucleotide + phosphate (NADP ) to 2-oxoglutarate and CO2 in the presence of isocitrate dehydrogenase (ICDH), with the formation of reduced nicotinamide-adenine dinucleotide phosphate (NADPH) (1). (ICDH) + + (1) D-Isocitric acid + NADP 2-oxoglutarate + CO2 + NADPH + H The amount of NADPH formed in this reaction is stoichiometric with the amount of D-isocitric acid. It is the NADPH which is measured by the increase in absorbance at 340 nm. Bound D-isocitric acid is released by alkaline hydrolysis (2), (3), and then measured using the same principle (1). (pH 9-10) (2) D-Isocitric acid ester + H2O D-isocitric acid + alcohol (pH 9-10) (3) D-Isocitric acid lactone + H2O D-isocitric acid SPECIFICITY, SENSITIVITY, LINEARITY AND PRECISION: The assay is specific for D-isocitric acid. D-malic acid, L-lactic acid, L-aspartic acid and fumaric acid do not react. The smallest differentiating absorbance for the assay is 0.005 absorbance units. This corresponds to 0.177 mg/L of sample solution at the maximum sample volume of 2.00 mL (or to 3.54 mg/L with a sample volume of 0.1 mL). -

Detection and Formation Scenario of Citric Acid, Pyruvic Acid, and Other Possible Metabolism Precursors in Carbonaceous Meteorites
Detection and formation scenario of citric acid, pyruvic acid, and other possible metabolism precursors in carbonaceous meteorites George Coopera,1, Chris Reeda, Dang Nguyena, Malika Cartera, and Yi Wangb aExobiology Branch, Space Science Division, National Aeronautics and Space Administration-Ames Research Center, Moffett Field, CA 94035; and bDevelopment, Planning, Research, and Analysis/ZymaX Forensics Isotope, 600 South Andreasen Drive, Suite B, Escondido, CA 92029 Edited by David Deamer, University of California, Santa Cruz, CA, and accepted by the Editorial Board July 1, 2011 (received for review April 12, 2011) Carbonaceous meteorites deliver a variety of organic compounds chained three-carbon (3C) pyruvic acid through the eight-carbon to Earth that may have played a role in the origin and/or evolution (8C) 7-oxooctanoic acid and the branched 6C acid, 3-methyl- of biochemical pathways. Some apparently ancient and critical 4-oxopentanoic acid (β-methyl levulinic acid), Fig. 1, Table S1. metabolic processes require several compounds, some of which 2-methyl-4-oxopenanoic acid (α-methyl levulinic acid) is tenta- are relatively labile such as keto acids. Therefore, a prebiotic setting tively identified (i.e., identified by mass spectral interpretation for any such individual process would have required either a only). As a group, these keto acids are relatively unusual in that continuous distant source for the entire suite of intact precursor the ketone carbon is located in a terminal-acetyl group rather molecules and/or an energetic and compact local synthesis, parti- than at the second carbon as in most of the more biologically cularly of the more fragile members. -

Profiling Taste and Aroma Compound Metabolism During Apricot Fruit Development and Ripening
Int. J. Mol. Sci. 2016, 17, 998; doi:10.3390/ijms17070998 S1 of S4 Supplementary Materials: Profiling Taste and Aroma Compound Metabolism during Apricot Fruit Development and Ripening Wanpeng Xi, Huiwen Zheng, Qiuyun Zhang and Wenhui Li Figure S1. Sugars HPLC chromatogram of apricot fruit. Peaks (1) Fructose (2) Glucose (3) Sucrose. (a) sugars mixture standards; (b) sugars for SG peel of S5; (c) sugars for SG pulp of S5. Int. J. Mol. Sci. 2016, 17, 998; doi:10.3390/ijms17070998 S2 of S4 Figure S2. Organic acids HPLC chromatogram of apricot fruit. Peaks (1) oxalic acid (2) tartaric acid (3) quininic acid (4) malic acid (5) citric acid (6) fumaric acid. (a) organic acids mixture standard; (b) organic acid for YC peel of S4; (c) organic acid for YC pulp of S4. Int. J. Mol. Sci. 2016, 17, 998; doi:10.3390/ijms17070998 S3 of S4 Table S1. Chroma values of apricot fruit during development and ripening. L* a* b* C* H Cultivars S1 S2 S3 S4 S5 S1 S2 S3 S4 S5 S1 S2 S3 S4 S5 S1 S2 S3 S4 S5 S1 S2 S3 S4 S5 DX 54.52 57.13 61.56 62.61 55.49 20.28 18.13 14.41 8.87 17.97 39.02 37.67 38.27 39.99 43.83 43.98 42.27 41.06 42.26 47.67 117.46 116.92 110.21 76.26 67.36 HY 54.97 55.83 61.53 65.08 65.11 −16.04 −14.77 −13.30 11.01 12.78 34.44 31.68 32.74 38.82 42.51 37.99 34.84 35.35 52.69 45.14 114.91 114.94 112.05 72.65 72.57 KE 47.02 47.00 48.32 60.11 60.66 −17.08 −17.37 −16.65 −0.36 8.20 30.83 29.09 29.08 48.23 48.48 35.26 33.88 33.51 48.33 49.33 118.94 120.86 119.85 90.67 118.02 AK 50.02 50.90 52.19 68.35 60.28 −21.98 −21.76 −19.02 -4.97 4.04 40.07 39.12 35.71 -

Challenges in Analysis of Hydrophilic Metabolites Using Chromatography Coupled with Mass Spectrometry
Journal of Analysis and Testing https://doi.org/10.1007/s41664-020-00126-z REVIEW Challenges in Analysis of Hydrophilic Metabolites Using Chromatography Coupled with Mass Spectrometry Qingyu Hu1,2,3 · Huiru Tang3 · Yulan Wang4 Received: 2 February 2020 / Accepted: 26 March 2020 © The Nonferrous Metals Society of China 2020 Abstract Hydrophilic metabolites play important roles in cellular energy metabolism, signal transduction, immunity. However, there are challenges in both identifcation and quantifcation of the hydrophilic metabolites due to their weak interactions with C18-reversed-phase liquid chromatography (RPLC), leading to poor retention of hydrophilic metabolites on the columns. Many strategies have been put forward to increase the retention behavior of hydrophilic metabolites in the RPLC system. Non- derivatization methods are mainly focused on the development of new chromatographic techniques with diferent separation mechanisms, such as capillary electrophoresis, ion-pairing RPLC etc. Derivatization methods improve the hydrophobicity of metabolites and can enhance the MS response. This review mainly focused on the illustration of challenges of LCMS in the analysis of hydrophilic metabolomics feld, and summarized the non-derivatization and derivatization strategies, with the intention of providing multiple choices for analysis of hydrophilic metabolites. Keywords Hydrophilic metabolites · Hydrophilic interaction chromatography · Ion-pairing reversed-phase liquid chromatography · Ion chromatography · Capillary electrophoresis · -

Continuous Citric Acid Secretion by a High Specific Ph Dependent Active Transport System in Yeast Candida Oleophila ATCC 20177
Electronic Journal of Biotechnology ISSN: 0717-3458 Vol.8 No.2, Issue of August 15, 2005 © 2005 by Pontificia Universidad Católica de Valparaíso -- Chile Received November 22, 2004 / Accepted March 28, 2005 RESEARCH ARTICLE Continuous citric acid secretion by a high specific pH dependent active transport system in yeast Candida oleophila ATCC 20177 Savas Anastassiadis*# Department of Environmental Engineering School of Engineering Democritus University of Thrace 67100 Xanthi, Greece E-mail: [email protected] Hans-Jürgen Rehm Institute of Microbiology University of Münster Corrensstr. 3, 48149 Münster, Germany (retired Professor) Website: http://www.greekbiotechnologycenter.gr Financial support: Part of the work that has been carried out at the Institute of Biotechnology 2 of Research Centre Jülich (Germany) was financed by Haarmann and Reimer, a daughter company of the company Bayer, Leverkusen, Germany. Keywords: active citrate export, citric acid fermentation, energy consuming citric acid secretion, specific active transport system. Present address: #Research in Biotechnology, Co., Vat. #: 108851559. Avgi/Sohos, 57002 Thessaloniki, Greece; Tel. +30-2395-051324; +30-6973- 801395 (cellular); Tel./Fax. +30-2395-051470, E-mail: [email protected]. The pH influence on continuous citric acid secretion was similarities between A. niger and yeast strains in investigated in Candida oleophila ATCC 20177 (var.) mechanism of citric acid synthesis, however, differences + under NH4 limiting state steady conditions, using still exist in terms of triggering out and regulation of citrate glucose. Highest citric acid concentration of 57.8 g/l, overproduction. Many models have been developed citrate/isocitrate ratio of 15.6, space-time yield of 0.96 describing the biochemistry of citrate synthesis, using g/(l x hr) and biomass specific productivity of 0.041 g/(g glucose and other carbon sources, however a complete x hr) were obtained at pH 5 and 60 hrs residence time. -

136757A0.Pdf
NOVEMBER 9, 1935 NATURE 757 quantitatively by the aspartase enzyme, and identified Using the ordinary technique, we have examined qualitatively in the form of the copper salt. Approxi specimens of purified l-fructose and l-arabinose, and mately the other half of the nitrogen ( 40-50 per cent) in neither event have we been able to detect the is precipitable by phosphotungstic acid. Since slightest reducing action on Tillmans's reagent. We cystine, arginine, histidine and aromatic amino acids have no explanation to offer of Jonnissian's results, are absent, it is probable that this fraction consists but we thought it might be of interest to publish of lycine. Further work is in progress to determine the above facts, in case that author's statements the nature of the compounds precipitated by phospho should lead other workers to under-value what has tungstic acid. come to be recognised as an extremely useful reagent. In an earlier paper1, it was mentioned that aspartic A. L. BACHARACH. acid is not present in the amino acid mixture. This H. E. GLYNN. erroneous conclusion is ascribable to the fact that, Glaxo Laboratories, Ltd., owing to the small amount of material available, the 56 Osnaburgh Street, determination was made from the solution from London, N.W.l. which ammonia had first been distilled off according Oct. 18. to van Slyke, in which process aspartic acid seems to • Proc. 6th Caw,. Cong. Phys. Pharm. Bio., Academy of Sciences be precipitated as a calcium salt. Press, Moscow and Leningrad, 101; 1935. The composition of the excreted amino acid mixture is very interesting in several respects. -

Citric Acid Cycle
Lecture: 4 Biochemistry Anwar J Almzaiel Citric acid cycle Citric acid cycle (Krebs cycle, tricarboxylic acid cycle) is a series of reactions in mitochondria that bring about the catabolism of acetyl residues, liberating hydrogen equivalents, which upon oxidation lead to the release of most the energy of tissues fuels. The major function of cycle is to act as the final common pathway for oxidation of fatty acids, carbohydrates and proteins. It includes the electron transport system, the energy is produced in large amounts. Several of these processes are carried out in many tissues but the liver is the only tissue in which all occur to a significant extent, thus it is lethal when large numbers of hepatic cell are damaged or replaced by connective tissue, as in acute hepatitis and cirrhosis respectively. This cycle is found in the mitochondria. It starts with pyruvic acid which comes from cytoplasm to the mitochondria to be converted with coenzymes (NAD) and enzymes into acetyl COA + Pyruvic acid + COA +NAD Acetyl CoA +NADH+ CO2 The enzymes needed are found in the mitochondria close to the enzymes of the respiratory chain The oxidation of pyruvate to acetyl-CoA Before pyruvate can enter the citric acid cycle, it must be transported into the mitochondria via a special pyruvate transporter that aids its passage across the inner mitochondrial membrane. Within the mitochondria, pyruvate is oxdatively decarboxylated into acetyl COA. The conversion of pyruvic acid to acetyl COA involves 5 types of reaction and each reaction is catalysed by different enzyme system these enzymes act as a multi enzyme system (complex). -

Strecker Degradation Products of Aspartic and Glutamic Acids and Their Amides
Czech J. Food Sci. Vol. 19, No. 2: 41–45 Strecker Degradation Products of Aspartic and Glutamic Acids and their Amides JAN RÖSSNER, JAN VELÍEK, FRANTIEK PUDIL and JIØÍ DAVÍDEK Institute of Chemical Technology Department of Food Chemistry and Analysis, Prague, Czech Republic Abstract RÖSSNER J., VELÍEK J., PUDIL F., DAVÍDEK J. (2001): Strecker degradation products of aspartic and glutamic acids and their amides. Czech J. Food Sci., 19: 4145. Aspartic and glutamic acids, asparagine and glutamine were oxidised with either potassium peroxodisulphate or glyoxal. Non- volatile products were derivatised and analysed by GC/FID and GC/MS. Volatile reaction products were isolated and analysed by the same methods. It was found that the degradation reactions of amino acids are complex. Amino acids are principally degraded via the corresponding α-keto acids to Strecker aldehydes (aspartic acid to oxalacetic and 3-oxopropionic acids and glutamic acid to α-ketoglutaric and 4-oxobutyric acids), which are unstable and decomposed by decarboxylation to the corresponding alde- hydes. Aspartic acid also eliminates ammonia and yields fumaric acid whereas glutamic acid gives rise to an imine, pyroglutamic acid. A recombination of free radicals leads to dicarboxylic acids (succinic acid from aspartic acid, succinic, glutaric and adipic acids from glutamic acid). The major volatile products (besides the aldehydes) are lower carboxylic acids (acetic acid from aspartic acid and propionic acid acid from glutamic acid) that can at least partly arise by radical reactions. In both quality and quantity terms, a higher amount of degradation products arises by oxidation of amino acids by peroxodisulphate. Keywords: Strecker degradation; Strecker aldehydes; amino acids; glyoxal; sodium peroxodisulphate; aspartic acid; glutamic acid; asparagine; glutamine; radicals The reaction of an α-amino acid with an oxidation re- asparagine (Asn) and glutamine (Gln). -
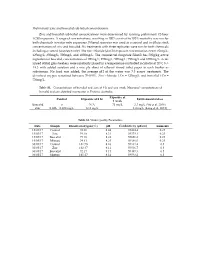
Supplementary File 1 (PDF, 650 Kib)
Preliminary Zinc and boscalid sub lethal concentrations Zinc and boscalid sub lethal concentrations were determined by running preliminary 72‐hour LC50 exposures. A range of concentrations, resulting in 100% survival to 100% mortality was run for both chemicals in water only exposures. Filtered seawater was used as a control and to dilute stock concentrations of zinc and boscalid. Six treatments with three replicates were run for both chemicals including a control (seawater only). The zinc chloride (ZnCl2) exposure concentrations were: 62mg/L; 125mg/L; 250mg/L; 500mg/L and 1000mg/L. The commercial fungicide Filan® has 500g/kg active ingredient of boscalid, concentrations of 100mg/L; 250mg/L; 500mg/L; 750mg/L and 1000mg/L. Acid‐ rinsed 600ml glass beakers were randomly placed in a temperature‐controlled incubator at 20oC (+/‐ 1oC) with added aeration and a one ply sheet of ethanol rinsed toilet paper in each beaker as substratum. No food was added; the average pH of the water was 7.5 across treatments. The dissolved oxygen remained between 70‐100%. Zinc chloride LC50 = 125mg/L and boscalid LC50 = 750mg/L. Table S1. –Concentrations of boscalid and zinc at 0 h and one week. Measured concentrations of boscalid and zinc detected in estuaries in Victoria, Australia. Exposure at Control Exposure at 0 hr Environmental dose 1 week Boscalid 0 N/A 75 mg/L 3.3 mg/L (Vu et al. 2016) Zinc 0.026 – 0.034 mg/L 12.5 mg/L 3.4 mg/L (Long et al. 2015) Table S2. Water Quality Parameters. Date Sample Dissolved oxygen (%) pH Conductivity (µS/cm) Ammonia 15/05/17 Control 98.36 8.44 59464.4 0.25 15/05/17 Zinc 99.10 8.31 59379.1 0.25 15/05/17 Boscalid 99.18 8.28 59656.8 0.25 15/05/17 Mixture 94.81 8.25 59386.0 0.25 30/05/17 Control 101.90 8.16 59332.4 0.5 30/05/17 Zinc 102.47 8.11 59536.7 0.5 30/05/17 Boscalid 92.17 8.15 59489.3 0.5 30/05/17 Mixture 101.57 8.14 59593.2 0.5 Supplementary Material .