Pianists Duet Better When They Play with Themselves: on the Possible Role of Action Simulation in Synchronization
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Comparative Analysis of the Six Duets for Violin and Viola by Michael Haydn and Wolfgang Amadeus Mozart
A COMPARATIVE ANALYSIS OF THE SIX DUETS FOR VIOLIN AND VIOLA BY MICHAEL HAYDN AND WOLFGANG AMADEUS MOZART by Euna Na Submitted to the faculty of the Jacobs School of Music in partial fulfillment of the requirements for the degree, Doctor of Music Indiana University May 2021 Accepted by the faculty of the Indiana University Jacobs School of Music, in partial fulfillment of the requirements for the degree Doctor of Music Doctoral Committee ______________________________________ Frank Samarotto, Research Director ______________________________________ Mark Kaplan, Chair ______________________________________ Emilio Colón ______________________________________ Kevork Mardirossian April 30, 2021 ii I dedicate this dissertation to the memory of my mentor Professor Ik-Hwan Bae, a devoted musician and educator. iii Table of Contents Table of Contents ............................................................................................................................ iv List of Examples .............................................................................................................................. v List of Tables .................................................................................................................................. vii Introduction ...................................................................................................................................... 1 Chapter 1: The Unaccompanied Instrumental Duet... ................................................................... 3 A General Overview -

Personality Traits and Music Experience Tracy A
University of South Florida Scholar Commons Graduate Theses and Dissertations Graduate School November 2017 Music Ensemble Participation: Personality Traits and Music Experience Tracy A. Torrance University of South Florida, [email protected] Follow this and additional works at: http://scholarcommons.usf.edu/etd Part of the Other Education Commons Scholar Commons Citation Torrance, Tracy A., "Music Ensemble Participation: Personality Traits and Music Experience" (2017). Graduate Theses and Dissertations. http://scholarcommons.usf.edu/etd/7100 This Dissertation is brought to you for free and open access by the Graduate School at Scholar Commons. It has been accepted for inclusion in Graduate Theses and Dissertations by an authorized administrator of Scholar Commons. For more information, please contact [email protected]. Music Ensemble Participation: Personality Traits and Music Experience by Tracy A. Torrance A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy in Music with a concentration in Music Education School of Music College of The Arts University of South Florida Major Professor: Jennifer A. Bugos, Ph.D. Darlene DeMarie, Ph.D. C. Victor Fung, Ph.D. William Hayden, D.A. David A. Williams, Ph.D. Date of Approval: November 15, 2017 Keywords: Music Psychology, Music Education, Big Five, Music Engagement Copyright © 2017, Tracy A. Torrance ACKNOWLEDGEMENTS There are a number of people I would like to thank who have travelled this doctoral path with me. First and foremost, I would like to thank my parents, Robert and Barbara Torrance, for their love, support, and belief in me. I would never have gotten this far in my academic career without them and the rest of my family. -

Music Ensemble
PDF generated on: 2021-09-23 16:08:36 AEST https://www.tasc.tas.gov.au/ Music Ensemble 5 LEVEL 2 TCE CREDIT POINTS COURSE CODE MSM205115 COURSE SPAN 2015 — 2019 COURSE STATUS CLOSED READING AND WRITING STANDARD NO MATHEMATICS STANDARD NO COMPUTERS AND INTERNET STANDARD NO Music Ensemble is a Level 2 course which requires participants to have the vocal or instrumental competence to play/perform as a member of an ensemble Instrumental skills for ensemble playing, Responding to Musical Direction, Rehearsal and Performance and Care and Safe Practice make up the four components of the course. Music Ensemble provides an extension opportunity for learners studying other music courses or for learners who wish to have their participation in a school based ensemble recognised. Rationale Practicing, rehearsing and presenting music as a member of an ensemble is an integral component of music education. Learners develop music literacy and performance skills, and learn to articulate their ideas through rehearsal discussions. Though not a requirement, performance of Australian compositions and arrangements are encouraged, including original compositions of ensemble members. A knowledge and understanding of Australian music enables learners to participate more effectively in, and contribute to, diverse aspects of Australian cultural life. Aims The collaborative learning style of Music Ensemble aims to engender skills of adaptability, creative thinking, and self-discipline through playing, using recognised repertoire and instrumentation, and developing skills and techniques appropriate to different genres and/or styles. Music Ensemble aims to foster resilience, cooperation and positive team membership. Once learned, these attributes may be applied to a range of music groups and to other contexts and environments such as the work place and community organisations. -

Composed for Six-Hands Piano Alti El Piyano Için Bestelenen
The Turkish Online Journal of Design, Art and Communication - TOJDAC ISSN: 2146-5193, April 2018 Volume 8 Issue 2, p. 340-363 THE FORM ANALYSIS OF “SKY” COMPOSED FOR SIX-HANDS PIANO Şirin AKBULUT DEMİRCİ Assoc. Prof. Education Faculty, Music Education Faculty. Uludag University, Turkey https://orcid.org/0000-0001-8904-4920 [email protected] Berkant GENÇKAL Assoc. Prof. State Conservatory, School for Music and Drama. Anadolu University, Turkey https://orcid.org/0000-0001-5792-2100 [email protected] ABSTRACT The piano, which is a solo instrument that takes part in educational process, can take role not only in instrumental education but also in chamber music education as well with the 6-hands pieces for three players that perform same composition on a single instrument. According to the international 6-hands piano literature, although the works of Alfred Schnittke's Hommage, Carl Czerny's Op.17 and 741, Paul Robinson's Pensees and Montmartre and many more are included, the Turkish piano literature has been found to have a limited number of compositions. The aim of the work is to contribute to the field by presenting musical analysis about the place and the importance of the educational use of the works composed for 6-hands which are extremely rare in the Turkish piano literature. As an example, the piece named Sky composed by Hasan Barış Gemici is considered. Analysis is supported by comparative methods in form, structuralism, rhythm, theoretical applications and performance; it also gives information about basic playing techniques. It is thought that this study carries importance in contributing to the limited number of Turkish 6-hands piano literature and researchers who will conduct research in this regard, in terms of creating resources for performers and composers who will interpret the literature. -

Download Program Notes
Notes on the Program By James M. Keller, Program Annotator, The Leni and Peter May Chair Piano Quintet in F minor, Op. 34 Johannes Brahms ohannes Brahms was 29 years old in 1862, seated at the other piano. Ironically, critics Jwhen he embarked on this seminal mas- now complained that the work lacked the terpiece of the chamber-music repertoire, sort of warmth that string instruments would though the work would not reach its final have provided — the opposite of Joachim’s form as his Piano Quintet until two years objection. Unlike the original string-quintet later. He was no beginner in chamber music version, which Brahms burned, the piano when he began this project. He had already duet was published — and is still performed written dozens of ensemble works before and appreciated — as his Op. 34bis. he dared to publish one, his B-major Piano By this time, however, Brahms must have Trio (Op. 8) of 1853–54. Among those early, grown convinced of the musical merits of his unpublished chamber pieces were 20-odd material and, with some coaxing from his string quartets, all of which he consigned friend Clara Schumann, he gave the piece to destruction prior to finally publishing his one more try, incorporating the most idiom- three mature works in that classic genre in atic aspects of both versions. The resulting the 1870s. In truth, he did get some use out Piano Quintet, the composer’s only essay of those early quartets — to paper the walls in that genre (and no wonder, after all that and ceilings of his apartment. -
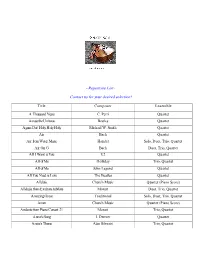
Repertoire List~
~Repertoire List~ Contact us for your desired selection! Title Composer Ensemble A Thousand Years C. Perri Quartet Across the Universe Beatles Quartet Agnus Dei/Holy Holy Holy Michael W. Smith Quartet Air Bach Quartet Air from Water Music Handel Solo, Duet, Trio, Quartet Air On G Bach Duet, Trio, Quartet All I Want is You U2 Quartet All of Me Holliday Trio, Quartet All of Me John Legend Quartet All You Need is Love The Beatles Quartet Alleluia Church Music Quartet (Piano Score) Allelujia from Exultate Jubilate Mozart Duet, Trio, Quartet Amazing Grace Traditional Solo, Duet, Trio, Quartet Amen Church Music Quartet (Piano Score) Andante from Piano Concert 21 Mozart Trio, Quartet Annie’s Song J. Denver Quartet Annie’s Theme Alan Silvestri Trio, Quartet Apollo 13 Horner Quartet Appalachia Waltz O’Connor Trio, Quartet Arioso Bach Solo, Duet, Trio, Quartet Arrival of the Queen of Sheba Handel Trio, Quartet Ashokan Farewell Jay Ungar Quartet (Piano Score) At Last Harry Warren Quartet Ave Maria Bach-Gounod Solo, Duet, Trio, Quartet Ave Maria (A flat) Schubert Quartet (Piano Score) Ave Maria (B flat) Schubert Quartet Ave Verum Corpus Mozart Solo, Duet, Trio, Quartet (plus Piano Score) Barcarolle from Tales of Hoffman Offenbach Quartet Be Thou My Vision Traditional Duet, Trio, Quartet Beauty and the Beast Ashman Quartet Bei Mennern from The Magic Flute Mozart Quartet Best Day of My life American Authors Quartet Bist du bei mer Bach Trio, Quartet Bittersweet Symphony M. Jagger Quartet Blessed are They Church Music Quartet Blessed are Those Who Love You Haugen Quartet (Piano Score) Blue Moon Rodgers Quartet Born Free Barry Quartet Bourree Bach Solo, Duet, Trio, Quartet Bourree Handel Solo, Duet, Trio, Quartet Brandenburg Concerto No. -

D Milhaud Scaramouche 1 Piano 4 Hands Sheet Music
D Milhaud Scaramouche 1 Piano 4 Hands Sheet Music Download d milhaud scaramouche 1 piano 4 hands sheet music pdf now available in our library. We give you 6 pages partial preview of d milhaud scaramouche 1 piano 4 hands sheet music that you can try for free. This music notes has been read 6326 times and last read at 2021-09-28 15:26:41. In order to continue read the entire sheet music of d milhaud scaramouche 1 piano 4 hands you need to signup, download music sheet notes in pdf format also available for offline reading. Instrument: 1 Piano 4 Hands Ensemble: Musical Ensemble Level: Advanced [ READ SHEET MUSIC ] Other Sheet Music D Milhaud Scaramouche Ii Modr For Alto Saxophone Solo String Quartet D Milhaud Scaramouche Ii Modr For Alto Saxophone Solo String Quartet sheet music has been read 2971 times. D milhaud scaramouche ii modr for alto saxophone solo string quartet arrangement is for Advanced level. The music notes has 6 preview and last read at 2021-09-27 03:30:57. [ Read More ] Scaramouche Scaramouche sheet music has been read 2058 times. Scaramouche arrangement is for Advanced level. The music notes has 1 preview and last read at 2021-09-27 12:27:39. [ Read More ] Arlequin Et Scaramouche Arlequin Et Scaramouche sheet music has been read 2102 times. Arlequin et scaramouche arrangement is for Advanced level. The music notes has 6 preview and last read at 2021-09-27 15:55:16. [ Read More ] Minimalist Tribute To Darius Milhaud For Piano Solo From Three Minimalist Tributes Minimalist Tribute To Darius Milhaud For Piano Solo From Three Minimalist Tributes sheet music has been read 4544 times. -

Broadway Musical Ensemble
School District of Springfield Township Springfield Township High School Course Overview Course Name: Broadway Musical Ensemble Course Description The Broadway Musical Ensemble class offers students the opportunity to produce and perform a Broadway musical each semester. Students learn the proper vocal techniques needed to perform a variety of musical theatre genres. The students also develop self-critiquing skills as they refine their talents for the performances. Students are required to perform for the public and are assessed on their class participation, written pre-production assignments, and final performance. Course Prerequisites Student audition and/or teacher recommendation Unit Titles Unit One: Introduce Musical Theatre Terminology, Choreography Skills, Vocal Skills, Script/Character Analysis; Write a one-act script based on a specific theme chosen by the class Unit Two: Set/Stage Direction Planning, Rehearsing Scenes, Dances and Musical Selections, Set Construction Work Unit Three: Dress Rehearsals, Performances and Formal Evaluations Essential Questions 1. What research is needed to understand the subject matter of the musical show? 2. What research does each performer need to do to understand his/her character, the plot/theme, and the relationships of the other characters? 3. What are the major terms and skills needed to direct, choreograph, and produce a musical show? 4. How does the performer astutely evaluate his/her dress rehearsal as well as the final performance? 5. What is the correct musical theatre terminology when directing and/or choreographing a scene or musical number? 6. How does the performer portray his/her character/part through the singing, acting and dancing? 7. How does one produce a final performance that is enlightening/educational as well as entertaining? 8. -

Music Appreciation April 16, 2020
Music Virtual Learning Music Appreciation April 16, 2020 Music Appreciation Lesson: April 16, 2020 Objective/Learning Target: Students will learn about musical characteristics of the classical period. Bell Work Look at this painting and think about our previous lesson on the social and artistic aspects of the classical era. Write about two aspects that you see present in this piece of art. Lesson Classical Period ● ● New Musical inventions: Classical Period ● ● ● ● ● ● Classical musical forms Symphony ● ● ● ● ● Orchestra performing on a stage Movement: a section of a musical work. Sonata ● ● Key: a group of notes that are used in a piece of music. Sonata Form I. Exposition-Contains two themes known as “subjects.” A. First Subject-In the Tonic Key or “Home key” B. Second Subject- Ends the section in the Dominant Key of “V” II. Development-develops both of those themes by using different key centers. End in the Tonic Key or “Home key.” III. Recapitulation- It’s a “Recap” of everything that’s happened. A. First and Second Subject return BOTH in the Tonic Key Ends with a “Coda”-a short passage that rounds out the music. Sonata Form Diagram Coda Exposition Development Recapitulation Trans. Coda 1st Trans. 2nd Coda in Develop 2nd 1st Link to (Tonic) Subject Link to Subject Dom. Themes Subject Subject next The (Tonic) next Dom. (V) (Tonic) Subject End! subject (V) Modulations Take a listen to… Beethoven: Symphony No. 5, 1st movement Concerto ● ● ● ● String Quartet String Quartet ● A string quartet is a musical ensemble of four string players – two violin players, a viola player and a cellist – or a piece written to be performed by such a group. -

Memorandum LIBRARY of CONGRESS
UNITED STATES GOVERNMENT Memorandum LIBRARY OF CONGRESS 5JSC/LC/12 TO: Joint Steering Committee for Development of RDA DATE: February 6, 2008 FROM: Barbara B. Tillett, LC Representative SUBJECT: Proposed revision of RDA chap. 6, Additional instructions for musical works and expressions The Library of Congress is submitting rule revision proposals for the RDA December 2007 draft chapter 6 instructions for musical works and expressions. Goals of proposals 1. To maintain the additional instructions for music intact (although LC recommends integrating them with the general instructions after the first release of RDA). 2. To fill in gaps in the AACR2 rules carried over into RDA by a. Incorporating selected AACR2 rule revisions; b. Adding instructions that clarify, make explicit, or expand some principles and instructions carried over from AACR2; c. Proposing new instructions. 3. To simplify some unnecessarily complex instructions. 4. To arrange certain subsections in the six major instructions in a more logical way, based wherever possible on principles that group together types of resources having common characteristics. 5. To revise the instructions for medium of performance to provide for all media found in resources and to do that using vocabulary that adheres as closely as possible to the principle of representation: incorporating words the composer or resource uses. 6. To revise or eliminate instructions LC finds unworkable based on past experience with AACR2 (e.g., the instructions for key (6.22) in section M of this document). 7. In the interest of simplification, to eliminate vexing terms catalogers have spent inordinate amounts of time interpreting when using AACR2 (e.g., “type of composition” as a formal term when all it need be is a useful phrase; “score order”). -

Music 130-205 Piano Ensemble
Music 130-205: Chamber Music Workshop Piano (Piano Ensemble) Syllabus – Spring 2013 Director: Pamela Cordle My Office: CAB-1038 Telephone: (910) 392-2721 Email: [email protected] Class Time: Tuesdays and Thursdays, 5:00-6:15 pm Class Location: CAB-1019 (piano lab) Class meets initially in the digital piano lab. We move into practice rooms as needed. Our Goal: We will explore, study, prepare, and successfully perform piano literature composed for one piano-four hands (duet), one piano-six hands (trio), two pianos-four hands (duo), and two pianos-eight hands (quartet). This course is designed to improve piano sight-reading skills, enhance musicianship, improve interpretive skills, and review good performance practice for piano music. The culmination of the semester is a performance in Beckwith Recital Hall on Sunday, April 14, 2013 at 7:30. Attendance policy: Attendance is required! Be on time at the beginning of each rehearsal and stay until the end. The success of the ensemble depends on quality practice time and efficient, effective rehearsal techniques by all members. Except in emergencies, absences will not be excused. Exceptions may be made in the case of contagious or extended illness, but only if you have notified me in advance of the absence. If you have to miss and wish to be excused, use the phone number listed above. Please leave a voice message explaining the reason for your absence. If you share music, deliver the score for your partner before rehearsal. If you have to miss class, you are responsible for asking for and practicing assignments you may have missed. -

6Th Grade Music: String Quartets
Name: ______________________________________ Music 6___________ 6th Grade Music: String Quartets Instructions: Read the information below (this came from the website https://www.liveabout.com/string-quartet-101-723913). Prompt #1: Answer these questions using complete sentences. 1. Use your own words to describe what a string quartet is. 2. String quartets are usually made up of four movements (sections). Which of the four movements are usually fast? Circle all that apply a. First movement b. Second movement c. Third movement d. Fourth movement. Prompt #2: Under “Notable String Quartet Composers” below, choose one YouTube video to listen to. You do not need to listen to the entire video. Answer the following questions as you watch and listen: 1. Write the name of the composer you chose. 2. Is the beginning slow (Adagio) or fast (Allegro)? 3. The musicians are seated in a certain order. How do their instruments relate to that order? Information was found on this website: https://www.liveabout.com/string-quartet-101-723913 4. How do the musicians respond to each other while playing? Prompt #3: Under Modern String Quartet Music, find the Love Story quartet by Taylor Swift. Listen to it as you answer using complete sentences. 1. Which instrument is plucking at the beginning of the piece? (If you forgot, violins are the smallest instrument, violas are a little bigger, cellos are bigger and lean on the floor). 2. Reflect on how this version of Love Story is different from Taylor Swift’s original song. How would you describe the differences? What does this version communicate? Information was found on this website: https://www.liveabout.com/string-quartet-101-723913 String Quartet 101 All You Need to Know About the String Quartet The Jerusalem Quartet, a string quartet made of members (from left) Alexander Pavlovsky, Sergei Bresler, Kyril Zlontnikov and Ori Kam, perform Brahms’s String Quartet in A minor at the 92nd Street Y on Saturday night, October 25, 2014.