The Effect of Helper Presence on Egg Size in Cooperatively Breeding Birds
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Species List
May 11 – 23, 2019 Spain: Birding and Nature Tour With: Christine, Laura, Brad, Cathy, Elizabeth, and Richard (HO)= Distinctive enough to be counted as heard only (I)=introduced Tour Summary: What an amazing destination! A journey through Spain is an experience one will never forget. Over the course of this tour, we explored wildlife rich areas from the lofty peaks of the Gredos Mountains to the mudflats of Andalusia, from the cork oak forests of Extremadura to the saline pools of Castilla-La Mancha. We recorded 188 bird species in that time – species such as Great Bustard, Egyptian Vulture, Eurasian Hoopoe, European Roller, Crested Tit, Great Spotted Cuckoo, Pin-tailed Sandgrouse, and more. In the end, there was no agreement as to the favorite bird of the tour – each person selected three completely different birds! It was just not the birds that captivated our group. It was the richness of Spain’s culture, history, architecture, and cuisine interwoven with the natural landscape. BIRDS (188 species recorded, 3 heard only): DUCKS, GEESE AND SWANS: Anatidae (10) Graylag Goose Anser anser—the ancestor of the domestic goose and quite common in winter in Spain, but only a small percentage remain to breed – a couple lingering birds at Dehesa Abajo in Andalusia and a family of adults and goslings at Laguna Navaseca in Castille-La Mancha Common Shelduck Tadorna tadorna—very handsome shelduck, associated with saline wetlands; our best views were at the various lagoons in Castille-La Mancha, where they were quite common Northern Shovler Spatula -

La Mancha, Coto Donana & Extremadura 2017
Field Guides Tour Report Spain: La Mancha, Coto Donana & Extremadura 2017 May 6, 2017 to May 18, 2017 Chris Benesh & Godfried Schreur For our tour description, itinerary, past triplists, dates, fees, and more, please VISIT OUR TOUR PAGE. Spectacular skies greeted us during our visit to old Trujillo in the heart of Extremadura. Photo by guide Chris Benesh. So many birds around that you don´t know which to choose and observe. Do you recognize this feeling? We experienced many of these exciting moments in Spain during the Field Guides tour in May. It started straight away, on the first day, overlooking the natural lagoons of La Mancha Húmeda, where we had the chance to observe a great variety of species of ducks, grebes, terns, and passerines. The highlights here were the White-headed Duck, Eared Grebe, Red-crested Pochard, Whiskered Tern and Penduline Tit. In the National Park of Coto Donana again we found ourselves surrounded by birds: larks, bee-eaters, flamingos, Great Reed Warblers, Glossy Ibis, Squacco and Purple herons and a surprisingly well showing Little Bittern. With a bit of searching, scanning and listening we were able to also detect Red-knobbed Coot, Marbled Teal and Isabelline (Western Olivaceous) Warbler. Later in the week, close to Trujillo (Extremadura), we all enjoyed the excursion on the open, rolling plains, with Great and Little bustards, Eurasian Roller, Hoopoe, Calandra Lark, Montagu´s Harrier and many, many White Storks. For the shy Black Storks we had to wait one day more. In Monfrague National Park we discovered 3 pairs nesting on the breathtaking cliff of Peña Falcón. -
ORL 5.1 Hypothetical Spp Final Draft01a.Xlsx
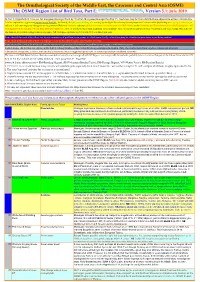
The Ornithological Society of the Middle East, the Caucasus and Central Asia (OSME) The OSME Region List of Bird Taxa, Part E: , Version 5.1: July 2019 In Part E, Hypothetical Taxa, we list non-passerines (prefixed by 'N') first, then passerines (prefixed by 'P'). Such taxa may be from distributions adjacent to or have extended to A fuller explanation is given in Explanation of the ORL, but briefly, Bright green shading of a row (eg Syrian Ostrich) indicates former presence of a taxon in the OSME Region. Light gold shading in column A indicates sequence change from the previous ORL issue. Red font indicates added information since the previous ORL version or the Conservation Threat Status (Critically Endangered = CE, Endangered = E, Vulnerable = V and Data Deficient = DD only). Not all synonyms have been examined. Serial numbers (SN) are merely an administrative convenience and may change. Please do not cite them in any formal correspondence or papers. NB: Compass cardinals (eg N = north, SE = southeast) are used. Rows shaded thus and with yellow text denote summaries of problem taxon groups in which some closely-related taxa may be of indeterminate status or are being studied. Rows shaded thus and with yellow text indicate recent or data-driven major conservation concerns. Rows shaded thus and with white text contain additional explanatory information on problem taxon groups as and when necessary. English names shaded thus are species on BirdLife Tracking Database, http://seabirdtracking.org/mapper/index.php. Only a few individuals from very few colonies are involved. A broad dark orange line, as below, indicates the last taxon in a new or suggested species split, or where sspp are best considered separately. -

Southern Spain: the Europe Introtour April 2019
Tropical Birding Trip Report Southern Spain: The Europe Introtour April 2019 A Tropical Birding set departure tour SOUTHERN SPAIN: The Europe Introtour 1st – 9th April 2019 Tour Leader: Emma Juxon All photographs in this report were taken by Emma Juxon unless otherwise stated, species depicted in photographs are named in BOLD RED www.tropicalbirding.com +1-409-515-9110 [email protected] Tropical Birding Trip Report Southern Spain: The Europe Introtour April 2019 Introduction For European birders, there is nowhere better to take a birding trip than the Iberian Peninsula. A region largely overlooked by North American birders in the past, it boasts some of THE best birding on the continent, it’s certainly not to be missed. We visit the outstanding regions of Extremadura and Andalucía on this tour, making our way through breathtaking mountainous landscapes, through rolling steppes and spectacular marismas. With many participants visiting the Old World for the first time, it promises a wealth of lifers, great food, fantastic people and an easy-going introduction to the Mediterranean way of life. One of the many beauties of this tour is that we only have two bases. Starting in Madrid, we make our way through the beautiful Spanish countryside, passing vineyards and castillos to get to wildflower- carpeted Extremadura. Here we enjoy the Belen Steppe, Caceres Plains and the exceptional Monfragüe National Park, encountering incredible birds such as Eurasian Griffon, Pin-tailed Sandgrouse, Great Bustard and Iberian Magpie. From here we head south to our next base in the picturesque pilgrimage town of El Rocío; I love this charming place, with its sandy roads, wonderful bird-filled marshes and charismatic people. -

Corvidae Species Tree
Corvidae I Red-billed Chough, Pyrrhocorax pyrrhocorax Pyrrhocoracinae =Pyrrhocorax Alpine Chough, Pyrrhocorax graculus Ratchet-tailed Treepie, Temnurus temnurus Temnurus Black Magpie, Platysmurus leucopterus Platysmurus Racket-tailed Treepie, Crypsirina temia Crypsirina Hooded Treepie, Crypsirina cucullata Rufous Treepie, Dendrocitta vagabunda Crypsirininae ?Sumatran Treepie, Dendrocitta occipitalis ?Bornean Treepie, Dendrocitta cinerascens Gray Treepie, Dendrocitta formosae Dendrocitta ?White-bellied Treepie, Dendrocitta leucogastra Collared Treepie, Dendrocitta frontalis ?Andaman Treepie, Dendrocitta bayleii ?Common Green-Magpie, Cissa chinensis ?Indochinese Green-Magpie, Cissa hypoleuca Cissa ?Bornean Green-Magpie, Cissa jefferyi ?Javan Green-Magpie, Cissa thalassina Cissinae ?Sri Lanka Blue-Magpie, Urocissa ornata ?White-winged Magpie, Urocissa whiteheadi Urocissa Red-billed Blue-Magpie, Urocissa erythroryncha Yellow-billed Blue-Magpie, Urocissa flavirostris Taiwan Blue-Magpie, Urocissa caerulea Azure-winged Magpie, Cyanopica cyanus Cyanopica Iberian Magpie, Cyanopica cooki Siberian Jay, Perisoreus infaustus Perisoreinae Sichuan Jay, Perisoreus internigrans Perisoreus Gray Jay, Perisoreus canadensis White-throated Jay, Cyanolyca mirabilis Dwarf Jay, Cyanolyca nanus Black-throated Jay, Cyanolyca pumilo Silvery-throated Jay, Cyanolyca argentigula Cyanolyca Azure-hooded Jay, Cyanolyca cucullata Beautiful Jay, Cyanolyca pulchra Black-collared Jay, Cyanolyca armillata Turquoise Jay, Cyanolyca turcosa White-collared Jay, Cyanolyca viridicyanus -

Seville and the Alentejo David Bradnum, Shaun Harvey & Howard Vaughan – June 2019
Seville and the Alentejo David Bradnum, Shaun Harvey & Howard Vaughan – June 2019 Overview and Logistics This was a short, low-cost birding break to Spain and Portugal. We had two aims: to see a handful of late-arriving spring migrants – White-rumped Swift, Western Olivaceous Warbler and Rufous-tailed Scrub-robin – and to enjoy as many as possible of the local specialities on the Castro Verde plains and the Rio Guadiana valley. We flew Easyjet from Gatwick to Seville. This cost c. £140 return each, booked just over a month in advance of the trip. We hired a car from Europcar, via Easyjet. This turned out to be a Skoda Octavia diesel for a bargain £65. Collection and drop-off were both relatively quick and fuss-free. We stayed in an Airbnb rental in the tiny Portuguese village of Bens, east of Mertola, for three nights. This was very convenient for the White-rumped Swift site and also another bargain, coming in at a total of just £130 for three nights! The traditional cottage was very comfortable and peaceful, with Iberian Magpies outside during the day and Red-necked Nightjar calling (once, at least!) during the night. We self-catered throughout, contributing to an overall cost of around £270 per person for the entire trip. The weather was generally good, with plenty of sunshine and peak temperature around 32°C (probably a little lower than might be expected). Day three was unusually overcast and even a little rainy around lunchtime – though this was actually helpful in that we could keep birding through the warmest part of the day! Ahead of the trip, we used the Finding Birds in Southern Portugal Gosney site guide to identify the best sites, and then topped this up with more recent info from eBird. -

Spain - Realm of the Iberian Lynx
Spain - Realm of the Iberian Lynx Naturetrek Tour Report 19 - 24 January 2020 Iberian Lynx Iberian Lynx Cinereous Vulture Sunset Report compiled by Niki Williamson Images by Simon Tonkin Naturetrek Mingledown Barn Wolf's Lane Chawton Alton Hampshire GU34 3HJ UK T: +44 (0)1962 733051 E: [email protected] W: www.naturetrek.co.uk Tour Report Spain - Realm of the Iberian Lynx Tour participants: Simon Tonkin & Niki Williamson (leaders) with 13 Naturetrek clients.. Summary For our band of patient nature-lovers, this six-day exploration of the realm of the Iberian Lynx gave us something amazing every day! Six different individuals of the endangered Spanish Imperial Eagle, rare Marbled Ducks, Hawfinches, Spanish Ibex cantering across a rock face, herds of Red Deer swimming a lake, duetting Little Owls, clouds of Cinereous and Griffon Vultures, Golden Eagles and shades of blue in the form of Bluethroat, Blue Rock Thrush, Iberian Magpie and Common Kingfisher were just some of our trip´s natural highlights. Our hosts´ hospitality was fantastic at both bases, and the group enjoyed sampling delicious local food such as chickpea and spinach stew, salmorejo soup and egg revuelto dishes, not to mention mouth-watering picnics in the sun, sometimes accompanied by dazzling flocks of Iberian Magpies, always accompanied by wine! Our fleeting glimpse of a female Iberian Lynx in Doñana Natural Park was to provide a suitable appetite-whetter for our superb encounter in Sierra de Morena, where a stunning female stalked across the track in front of us before taking up a pose on a nearby rock, allowing us to watch for over an hour! Day 1 Sunday 19th January Leaders Simon and Niki met the group as they converged on Sevilla airport, from various flights and pre-trip stays. -

Grey Shrikes Unless Noted Otherwise
Trends in systematics Speciation in shades of grey: Grey Shrike L elegans, while other great grey shrike taxa were left undetermined for the time being. the great grey shrike complex The purpose of this short paper is to present an Sometimes clear-cut species limits are hard to update on geographic variation in the great grey come by. A number of widespread Palearctic spe- shrike complex based on recent genetic studies cies and species complexes display an intricate (Gonzalez et al 2008, Klassert et al 2008, Olsson pattern of geographical (plumage) variation. et al 2010) and to show current implications for Information on patterns of genetic variation can species limits within this complex. Olsson et al be a tremendous help in clarifying relationships (2010) sampled by far the most extensively and between populations but the results are not al- agree with Gonzalez et al (2008) and Klassert et al ways unambiguous. The great grey shrike complex (2008) on the basic structure of the phylogeny. is one such diffcult case. Many may have been Therefore, Olsson et al (2010) is referred to below, surprised to note the treatment of great grey shrikes unless noted otherwise. in the second English edition of the Collins bird guide (Svensson et al 2009) in which two species Results are recognized: Great Grey Shrike Lanius excubi- The recovered mitochondrial DNA (mtDNA) tree tor and Iberian Grey Shrike L meridionalis. The lat- (fgure 1) shows a deep split between two large ter now only includes the birds from Iberia and clades, representing up to several million years of south-eastern France. -

Spain - Realm of the Iberian Lynx
Spain - Realm of the Iberian Lynx Naturetrek Tour Report 23 - 28 October 2017 Iberian Lynx by Stephen Debenham Iberian Lynx by Penny Fleming Iberian Grey Shrike by Penny Fleming Iberian Magpies by Penny Fleming Report compiled by Niki Williamson & Simon Tonkin Images courtesy of Penny Fleming and Stephen Debenham Naturetrek Mingledown Barn Wolf's Lane Chawton Alton Hampshire GU34 3HJ UK T: +44 (0)1962 733051 E: [email protected] W: www.naturetrek.co.uk Tour Report Spain - Realm of the Iberian Lynx Tour participants: Simon Tonkin and Niki Williamson (leaders) and a group of 11 Naturetrek clients Summary This two-base tour took in the atmospheric misty woodlands and wetlands of Doñana Natural Park, as well as the sweeping cork oak forests of the Sierra Morena near Andùjar. The group were thrilled by close encounters with Wild Boar, rutting Red and Fallow Deer, Egyptian Mongoose, Iberian Hare and two species of bats, as well as an impressive array of mountain, wetland and farmland birds, including Blue Rock Thrush, Azure-winged Magpies, Eurasian Hoopoe, Southern Grey Shrike, Cinereous Vulture, Crag Martin, Black-winged Stilts, Eurasian Spoonbills, and European Eagle-Owl. But of course, the outstanding highlight was our intense, prolonged experience with Fraile, a nine-year-old male Iberian Lynx, with whom we shared enthralling times at dawn and dusk as he rested and preened between bouts of devouring a recently killed Red Deer. Day 1 Monday 23rd October Simon and Niki met with Venetia, who had been enjoying a stay in Seville for a couple of days prior to the trip, at the airport. -

The Birdmen of the Pleistocene: on the Relationship Between Neanderthals and Scavenging Birds
Quaternary International xxx (2016) 1e7 Contents lists available at ScienceDirect Quaternary International journal homepage: www.elsevier.com/locate/quaint The birdmen of the Pleistocene: On the relationship between Neanderthals and scavenging birds * Stewart Finlayson a, b, , Clive Finlayson b, c a Department of Life Sciences, Anglia Ruskin University, East Road, Cambridge, Cambridgeshire CB1 1PT, United Kingdom b Department of Natural History, The Gibraltar Museum, 18-20 Bomb House Lane, Gibraltar c Institute of Life and Earth Sciences, The University of Gibraltar, Gibraltar Museum Associate Campus, 18-20 Bomb House Lane, Gibraltar article info abstract Article history: We have examined 192 Middle Palaeolithic sites in the Palaearctic which have raptor and corvid bones Available online xxx associated in human occupation contexts. We have also examined 395 sites with Upper Palaeolithic contexts for comparison. We show that Neanderthals were regularly associated with a suite of birds of Keywords: prey and corvids. We identify that the main species were regular or seasonal scavengers which co- Neanderthals occurred across large areas of the Neanderthal geographical range. This suggests a long-standing in- Modern humans ter-relationship between Neanderthals, raptors and corvids. We propose that the degree of difficulty of Raptors capturing these species was not an insurmountable problem for the Neanderthals and provide present- Corvids Scavengers day examples of close interaction between scavenging birds and people. We also show that modern Mid-latitude belt humans had a similar relationship with the same suite of birds as the Neanderthals. We suggest that one possibility is that Neanderthals transmitted the behaviour to modern humans. © 2016 Elsevier Ltd and INQUA. -

Birding Tour
2021 BULGARIA APRIL 25TH – MAY 9TH, 2021 [Birding Tour]; BULGARIA ‘Classic Spring Birding’ The 2018 Tour recorded 232 species; expect up to 22 warblers, 9 tits, 10 woodpeckers, 5 owls, and 24 raptors, vultures, and falcons. Rare migrant birds mostly arrive from East Africa or West Asia. Food is delightful, travel is easy, accommodations are cozy, and dozens of times each day the scenic countryside shouts ‘photo’. Tour Style [B] $3,945/pp MEXICO JUNE 5TH - 19TH, 2021 [Birding Tour] NORTHERN MEXICO’S SUMMER ENDEMICS Breeding endemics of Chihuahua's Sierra Madre Occidental, Saltillo Grasslands, Sierra Madre Oriental, and Tamaulipan coastal plain for Thicket Tinamou, Thick-billed and Maroon-fronted parrots, Yellow- headed and Red-crowned amazons, Eared Quetzal, Tamaulipas Pygmy- Owl, Tawny-collared Nightjar, Colima Warbler, Altamira Yellowthroat, Crimson-collared Grosbeak, and Worthen’s Sparrow. Tour Style [C] $3,995/pp CHILE OCTOBER 27TH - NOVEMBER 19TH, 2021 NORTHERN CHILE - The Atacama Desert and the Chilean Altiplan0 Amid an arid and rugged landscape with the least amount of rainfall on earth and high-elevation alkaline lakes... 'Good Heavens, what are we doing here?' Why birds of course --James's Flamingo, Giant Coot, Peruvian Thick-knee, Inca Tern, and Chilean Woodstar. CENTRAL CHILE - The towering Andes and the Humboldt Current Vineyards, farmland, rugged mountains; the lower slopes hold Chilean Tinamou, Moustached Turca, Dusky Tapaculo, Crag Chilia, Patagonian Forest Earthcreeper, and Dusky-tailed Canastero. The highlands possess Diademed Sandpiper-Plover, and Ochre-naped, Rufous-naped, White-browed and Black-fronted ground-tyrants. Major highlights of the pelagic trip include up to six species of albatrosses. -

Spain: the Best of Europe a Tropical Birding Set Departure
Spain: The Best of Europe A Tropical Birding Set Departure March 22—28, 2015 Guides: Ken Behrens and Scott Watson Text by Ken Behrens Photos by Ken Behrens unless noted otherwise TOUR SUMMARY Spain is perhaps Europe’s best-known birding destination, and for good reason. It offers some of the continent’s finest birding, on top of charming and historic towns, beautiful landscapes, and wonderful food. For someone looking to make a first birding trip to Europe, Spain is an obvious choice. You can rack up a large proportion of the continent’s birdlife in a single short trip. For this reason, we consider this trip a sort of “Europe Introtour”. This short Tropical Birding trip takes in some of Spain’s most storied birding destinations: Monfragüe National Park and the semi-steppes around Trujillo, in Extremadura; and the famous Coto Doñana National Park and surrounds in Andalucía. Despite being only seven days long, we racked up 184 bird species! One of the great things about this tour is that we are based in only two hotels, for three nights each. One hotel is a beautiful and castle-like structure set in a vineyard, while the other is on the shores of the Doñana wetlands, with birding literally on the doorstep. Another great thing about this tour is that it is short, and that Spain can be reached by a relatively short cross-Atlantic flight, making it accessible for people with limited vacation time, or limited willingness to take long-haul flights. For those interested in a longer, two-week trip, this Spain trip Spain Set Departure Tour Mar.