Volatile-Organic Compound Changes in Rose Twigs Consequent to Infection with Rose Powdery Mildew
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Confinement Strategy to Prepare N-Doped Reduced Graphene Oxide
Electronic Supplementary Material (ESI) for Nanoscale. This journal is © The Royal Society of Chemistry 2019 Supporting Information A confinement strategy to prepare N-doped reduced graphene oxide foams with desired monolithic structures for supercapacitors Daoqing Liu,a,b,# Qianwei Li, *c,# Si Li, a,b Jinbao Hou,d Huazhang Zhao *a,b a Department of Environmental Engineering, Peking University, Beijing 100871, China b The Key Laboratory of Water and Sediment Sciences, Ministry of Education, Beijing 100871, China c State Key Laboratory of Heavy Oil Processing, Beijing Key Laboratory of Oil and Gas Pollution Control, China University of Petroleum, 18 Fuxue Road, Changping District, Beijing 102249, China d College of Chemical Engineering, Beijing University of Chemical Technology, Beijing 100029, China # Co-first author. * Corresponding author. Tel/fax: +86-10-62758748; email address: [email protected] (H.Z. Zhao). S1 Experimental Methods 1. Preparation of graphite oxide (GO) Graphite oxide was prepared from natural graphite (Jinglong Co., Beijing, China) according to 1 a modified Hummers method : 120 mL 98 wt% H2SO4 was poured into a beaker containing a mixture of 5 g natural graphite and 2.5 g NaNO3, and then the mixture was stirred in an ice bath for 30 min. 15 g KMnO4 was added slowly into the mixture, which was allowed to react for 2 h at a temperature no more than 20°C. Then, the temperature was risen to 35°C, and the reaction was performed for another 2 h. After that, the reactant mixture was poured slowly into 360 mL distilled water under violent stirring condition so as to control the temperature no more than 90°C, followed by further reaction at 75°C for 1 h. -

Sections-A, B and C
Science IX Sample Paper 7 Solved www.rava.org.in CLASS IX (2019-20) SCIENCE (CODE 086) SAMPLE PAPER-7 Time : 3 Hours Maximum Marks : 80 General Instructions : (i) The question paper comprises of three sections-A, B and C. Attempt all the sections. (ii) All questions are compulsory. (iii) Internal choice is given in each sections. (iv) All questions in Section A are one-mark questions comprising MCQ, VSA type and assertion-reason type questions. They are to be answered in one word or in one sentence. (v) All questions in Section B are three-mark, short-answer type questions. These are to be answered in about 50-60 words each. (vi) All questions in Section C are five-mark, long-answer type questions. These are to be answered in about 80-90 words each. (vii) This question paper consists of a total of 30 questions. 6. Who proposed the fluid mosaic model of protoplasm? [1] Section - A (a) Singer and Nicolson (b) Watson and Crick (c) Robert Hook (d) Robert Brown 1. Which of the following actions a force can do? [1] (a) Can move a stationary object. Ans : (a) Singer and Nicolson. (b) Can stop a moving object. or (c) Can change the speed of a moving object. Which of the following are complex tissues? (d) All of the above. (a) Xylem and Phloem Ans : (d) All of the above (b) Collenchyma and Sclerenchyma (c) Parenchyma and Collenchyma 2. Ozone layer protects us from which one of the (d) Xylem and Parenchyma following? [1] (a) X- rays. -

QUINOA Botany, Production and Uses Dedicated to the Loving Memory of My Parents, Who Departed for the Heavenly Abode on 16 January 2001
QUINOA Botany, Production and Uses Dedicated to the loving memory of my parents, who departed for the heavenly abode on 16 January 2001 Atul Bhargava QUINOA Botany, Production and Uses Atul Bhargava and Shilpi Srivastava CABI is a trading name of CAB International CABI CABI Nosworthy Way 38 Chauncey Street Wallingford Suite 1002 Oxfordshire OX10 8DE Boston, MA 02111 UK USA Tel: +44 (0)1491 832111 Tel: +1 800 552 3083 (toll free) Fax: +44 (0)1491 833508 Tel: +1 (0)617 395 4051 E-mail: [email protected] E-mail: [email protected] Website: www.cabi.org © A. Bhargava and S. Srivastava 2013. All rights reserved. No part of this publication may be reproduced in any form or by any means, electronically, mechanically, by photocopying, recording or otherwise, without the prior permission of the copyright owners. A catalogue record for this book is available from the British Library, London, UK. Library of Congress Cataloging-in-Publication Data Bhargava, Atul, 1975- Quinoa : botany, production and uses / Atul Bhargava, Shilpi Srivastava. p. cm. Includes bibliographical references and index. ISBN 978-1-78064-226-0 (alk. paper) 1. Quinoa. I. Srivastava, Shilpi. II. Title. SB177.Q55B43 2013 664′.7--dc23 2013009503 ISBN-13: 978 1 78064 226 0 Commissioning editor: Sreepat Jain Editorial assistant: Alexandra Lainsbury Production editor: Simon Hill Typeset by SPi, Pondicherry, India Printed and bound in the UK by CPI Group (UK) Ltd, Croydon, CR0 4YY. Contents Contributors vii Preface ix Acknowledgements xiii PART I – INTRODUCTION AND HISTORY 1. Introduction 1 2. Historical Perspectives and Domestication 16 Didier Bazile, Francisco Fuentes and Ángel Mujica 3. -

Eurysacca Melanocampta MEYRICK (LEPIDOPTERA: GELECHIIDAE) for TWO VARIETIES of QUINOA (Chenopodium Quinoa WILLD.) in OLFACTOMETRY ASSAYS
RESEARCH 71 PREFERENCE OF QUINOA MOTH: Eurysacca melanocampta MEYRICK (LEPIDOPTERA: GELECHIIDAE) FOR TWO VARIETIES OF QUINOA (Chenopodium quinoa WILLD.) IN OLFACTOMETRY ASSAYS Juan F. Costa1*, Walter Cosio1, Maritza Cardenas1, Erick Yábar1, and Ernesto Gianoli2 ABSTRACT Insects are attracted to plants by visual and olfactory cues. The quinoa moth, Eurysacca melanocampta Meyrick (Lepidoptera: Gelechiidae), is the main insect pest of the quinoa crop, Chenopodium quinoa Willd. (Chenopodiales: Chenopodiaceae), in the southern Peruvian Andes, causing grain yield losses. The aim of this study was to investigate the behavioural response of adult quinoa moths to olfactory stimuli. Specifically, the objectives of this study were: 1) to determine the capacity of E. melanocampta adults of searching for quinoa plants using plant olfactory cues; 2) to determine the preference of E. melanocampta females for the odours derived from two varieties of quinoa: Amarilla de Marangani and Blanca de Junín; and 3) to assess the attraction of male quinoa moths to E. melanocampta females and the host plant in olfactometric bioassays. Adults preferred quinoa plant odour sources in choice tests when distilled water was used as a control (P < 0.0001). Females were more attracted to the Blanca de Junín variety than to Amarilla de Marangani variety (P < 0.05). Males were more attracted to the odour derived from females than to the volatile compounds from plants (both varieties) or to the odour blend derived from plants plus females together. The level of attraction of males towards females is negatively affected by the presence of the quinoa plants. Key words: attraction, Chenopodiaceae, Gelechiidae, searching, olfactometry, quinoa. -

IOBC/WPRS Working Group “Integrated Plant Protection in Fruit
IOBC/WPRS Working Group “Integrated Plant Protection in Fruit Crops” Subgroup “Soft Fruits” Proceedings of Workshop on Integrated Soft Fruit Production East Malling (United Kingdom) 24-27 September 2007 Editors Ch. Linder & J.V. Cross IOBC/WPRS Bulletin Bulletin OILB/SROP Vol. 39, 2008 The content of the contributions is in the responsibility of the authors The IOBC/WPRS Bulletin is published by the International Organization for Biological and Integrated Control of Noxious Animals and Plants, West Palearctic Regional Section (IOBC/WPRS) Le Bulletin OILB/SROP est publié par l‘Organisation Internationale de Lutte Biologique et Intégrée contre les Animaux et les Plantes Nuisibles, section Regionale Ouest Paléarctique (OILB/SROP) Copyright: IOBC/WPRS 2008 The Publication Commission of the IOBC/WPRS: Horst Bathon Luc Tirry Julius Kuehn Institute (JKI), Federal University of Gent Research Centre for Cultivated Plants Laboratory of Agrozoology Institute for Biological Control Department of Crop Protection Heinrichstr. 243 Coupure Links 653 D-64287 Darmstadt (Germany) B-9000 Gent (Belgium) Tel +49 6151 407-225, Fax +49 6151 407-290 Tel +32-9-2646152, Fax +32-9-2646239 e-mail: [email protected] e-mail: [email protected] Address General Secretariat: Dr. Philippe C. Nicot INRA – Unité de Pathologie Végétale Domaine St Maurice - B.P. 94 F-84143 Montfavet Cedex (France) ISBN 978-92-9067-213-5 http://www.iobc-wprs.org Integrated Plant Protection in Soft Fruits IOBC/wprs Bulletin 39, 2008 Contents Development of semiochemical attractants, lures and traps for raspberry beetle, Byturus tomentosus at SCRI; from fundamental chemical ecology to testing IPM tools with growers. -

Measurements of Higher Alkanes Using NO Chemical Ionization in PTR-Tof-MS
Atmos. Chem. Phys., 20, 14123–14138, 2020 https://doi.org/10.5194/acp-20-14123-2020 © Author(s) 2020. This work is distributed under the Creative Commons Attribution 4.0 License. Measurements of higher alkanes using NOC chemical ionization in PTR-ToF-MS: important contributions of higher alkanes to secondary organic aerosols in China Chaomin Wang1,2, Bin Yuan1,2, Caihong Wu1,2, Sihang Wang1,2, Jipeng Qi1,2, Baolin Wang3, Zelong Wang1,2, Weiwei Hu4, Wei Chen4, Chenshuo Ye5, Wenjie Wang5, Yele Sun6, Chen Wang3, Shan Huang1,2, Wei Song4, Xinming Wang4, Suxia Yang1,2, Shenyang Zhang1,2, Wanyun Xu7, Nan Ma1,2, Zhanyi Zhang1,2, Bin Jiang1,2, Hang Su8, Yafang Cheng8, Xuemei Wang1,2, and Min Shao1,2 1Institute for Environmental and Climate Research, Jinan University, 511443 Guangzhou, China 2Guangdong-Hongkong-Macau Joint Laboratory of Collaborative Innovation for Environmental Quality, 511443 Guangzhou, China 3School of Environmental Science and Engineering, Qilu University of Technology (Shandong Academy of Sciences), 250353 Jinan, China 4State Key Laboratory of Organic Geochemistry and Guangdong Key Laboratory of Environmental Protection and Resources Utilization, Guangzhou Institute of Geochemistry, Chinese Academy of Sciences, 510640 Guangzhou, China 5State Joint Key Laboratory of Environmental Simulation and Pollution Control, College of Environmental Sciences and Engineering, Peking University, 100871 Beijing, China 6State Key Laboratory of Atmospheric Boundary Physics and Atmospheric Chemistry, Institute of Atmospheric Physics, Chinese -

Combining Applied and Basic Research
Understanding the Powdery Mildew Disease of the Ornamental Plant Phlox: Combining Applied and Basic Research Dissertation Presented in Partial Fulfillment of the Requirements for the Degree Doctor of Philosophy in the Graduate School of The Ohio State University By Coralie Farinas Graduate Program in Plant Pathology The Ohio State University 2020 Dissertation Committee Dr. Francesca Peduto Hand, Advisor Dr. Pablo S. Jourdan Dr. Thomas K. Mitchell Dr. Pierce A. Paul Dr. Jason C. Slot Copyrighted by Coralie Farinas 2020 1 Abstract The characterization of plant germplasm has tremendous potential to help address the many challenges that the field of plant health is facing, such as climate change continuously modifying the regions of previously known disease occurrence. The worldwide trade of the plant genus Phlox represents an important revenue for the horticultural industry. However, Phlox species are highly susceptible to the fungal disease powdery mildew (PM), and infected materials shipping across borders accelerate the risk of disease spread. Through collaboration with laboratories in the U.S., we investigated the genotypic and phenotypic diversity of a PM population to better understand its capacity to adapt to new environments and new resistant hosts. To do this, we developed tools to grow and study PM pathogens of Phlox in vitro, and then used whole genome comparison and multilocus sequence typing (MLST) analysis to study the genetic structure of the population. Additionally, we explored Phlox germplasm diversity to identify a range of plant responses to PM infection by comparing disease severity progression and length of latency period of spore production across a combination of Phlox species and PM isolates in vitro. -
Supporting Information for Modeling the Formation and Composition Of
Supporting Information for Modeling the Formation and Composition of Secondary Organic Aerosol from Diesel Exhaust Using Parameterized and Semi-explicit Chemistry and Thermodynamic Models Sailaja Eluri1, Christopher D. Cappa2, Beth Friedman3, Delphine K. Farmer3, and Shantanu H. Jathar1 1 Department of Mechanical Engineering, Colorado State University, Fort Collins, CO, USA, 80523 2 Department of Civil and Environmental Engineering, University of California Davis, Davis, CA, USA, 95616 3 Department of Chemistry, Colorado State University, Fort Collins, CO, USA, 80523 Correspondence to: Shantanu H. Jathar ([email protected]) Table S1: Mass speciation and kOH for VOC emissions profile #3161 3 -1 - Species Name kOH (cm molecules s Mass Percent (%) 1) (1-methylpropyl) benzene 8.50×10'() 0.023 (2-methylpropyl) benzene 8.71×10'() 0.060 1,2,3-trimethylbenzene 3.27×10'(( 0.056 1,2,4-trimethylbenzene 3.25×10'(( 0.246 1,2-diethylbenzene 8.11×10'() 0.042 1,2-propadiene 9.82×10'() 0.218 1,3,5-trimethylbenzene 5.67×10'(( 0.088 1,3-butadiene 6.66×10'(( 0.088 1-butene 3.14×10'(( 0.311 1-methyl-2-ethylbenzene 7.44×10'() 0.065 1-methyl-3-ethylbenzene 1.39×10'(( 0.116 1-pentene 3.14×10'(( 0.148 2,2,4-trimethylpentane 3.34×10'() 0.139 2,2-dimethylbutane 2.23×10'() 0.028 2,3,4-trimethylpentane 6.60×10'() 0.009 2,3-dimethyl-1-butene 5.38×10'(( 0.014 2,3-dimethylhexane 8.55×10'() 0.005 2,3-dimethylpentane 7.14×10'() 0.032 2,4-dimethylhexane 8.55×10'() 0.019 2,4-dimethylpentane 4.77×10'() 0.009 2-methylheptane 8.28×10'() 0.028 2-methylhexane 6.86×10'() -

View the PDF File of the Tachinid Times, Issue 14
The Tachinid Times ISSUE 14 February 2001 Jim O'Hara, editor Agriculture & Agri-Food Canada, Systematic Entomology Section Eastern Cereal and Oilseed Research Centre C.E.F., Ottawa, Ontario, Canada, K1A 0C6 Correspondence: [email protected] A couple of significant changes to The Tachinid the newsletter before the end of next January. This Times have taken place this year. Firstly, the newsletter newsletter appears first in hardcopy and then on the Web has moved to a new location: http://res2.agr.ca/ecorc/ some weeks later. isbi/tachinid/times/index.htm. Secondly, it is being produced as an Acrobat® PDF (Portable Document Format) Study on the phylogeny and diversity of Higher file for the first time. Though this format may be Diptera in the Northern Hemisphere (by H. Shima) inconvenient for some readers, it has a number of In 1999 I applied to the Japanese Government compelling advantages. It allows me to produce the (Ministry of Education, Science, Culture and Sports) for newsletter faster because there is a one-step conversion a 3-year research grant to fund an international project from a WordPerfect® file with embedded colour images under the general title, "Study on the Phylogeny and to a PDF file. Also, the result is a product that can be Diversity of Higher Diptera in the Northern Hemi- viewed on the Web (using the free Acrobat® Reader that sphere." Funding was approved in 2000 and the first is readily available online), downloaded from the Web, or meeting of the project team was held in Fukuoka at the distributed in hardcopy – each with exactly the same Biosystematics Laboratory of Kyushu University in late pagination and appearance. -

Vapor Pressures and Vaporization Enthalpies of the N-Alkanes from 2 C21 to C30 at T ) 298.15 K by Correlation Gas Chromatography
BATCH: je1a04 USER: jeh69 DIV: @xyv04/data1/CLS_pj/GRP_je/JOB_i01/DIV_je0301747 DATE: October 17, 2003 1 Vapor Pressures and Vaporization Enthalpies of the n-Alkanes from 2 C21 to C30 at T ) 298.15 K by Correlation Gas Chromatography 3 James S. Chickos* and William Hanshaw 4 Department of Chemistry and Biochemistry, University of MissourisSt. Louis, St. Louis, Missouri 63121 5 6 The temperature dependence of gas chromatographic retention times for n-heptadecane to n-triacontane 7 is reported. These data are used to evaluate the vaporization enthalpies of these compounds at T ) 298.15 8 K, and a protocol is described that provides vapor pressures of these n-alkanes from T ) 298.15 to 575 9 K. The vapor pressure and vaporization enthalpy results obtained are compared with existing literature 10 data where possible and found to be internally consistent. Sublimation enthalpies for n-C17 to n-C30 are 11 calculated by combining vaporization enthalpies with fusion enthalpies and are compared when possible 12 to direct measurements. 13 14 Introduction 15 The n-alkanes serve as excellent standards for the 16 measurement of vaporization enthalpies of hydrocarbons.1,2 17 Recently, the vaporization enthalpies of the n-alkanes 18 reported in the literature were examined and experimental 19 values were selected on the basis of how well their 20 vaporization enthalpies correlated with their enthalpies of 21 transfer from solution to the gas phase as measured by gas 22 chromatography.3 A plot of the vaporization enthalpies of 23 the n-alkanes as a function of the number of carbon atoms 24 is given in Figure 1. -
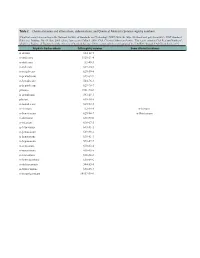
Table 2. Chemical Names and Alternatives, Abbreviations, and Chemical Abstracts Service Registry Numbers
Table 2. Chemical names and alternatives, abbreviations, and Chemical Abstracts Service registry numbers. [Final list compiled according to the National Institute of Standards and Technology (NIST) Web site (http://webbook.nist.gov/chemistry/); NIST Standard Reference Database No. 69, June 2005 release, last accessed May 9, 2008. CAS, Chemical Abstracts Service. This report contains CAS Registry Numbers®, which is a Registered Trademark of the American Chemical Society. CAS recommends the verification of the CASRNs through CAS Client ServicesSM] Aliphatic hydrocarbons CAS registry number Some alternative names n-decane 124-18-5 n-undecane 1120-21-4 n-dodecane 112-40-3 n-tridecane 629-50-5 n-tetradecane 629-59-4 n-pentadecane 629-62-9 n-hexadecane 544-76-3 n-heptadecane 629-78-7 pristane 1921-70-6 n-octadecane 593-45-3 phytane 638-36-8 n-nonadecane 629-92-5 n-eicosane 112-95-8 n-Icosane n-heneicosane 629-94-7 n-Henicosane n-docosane 629-97-0 n-tricosane 638-67-5 n-tetracosane 643-31-1 n-pentacosane 629-99-2 n-hexacosane 630-01-3 n-heptacosane 593-49-7 n-octacosane 630-02-4 n-nonacosane 630-03-5 n-triacontane 638-68-6 n-hentriacontane 630-04-6 n-dotriacontane 544-85-4 n-tritriacontane 630-05-7 n-tetratriacontane 14167-59-0 Table 2. Chemical names and alternatives, abbreviations, and Chemical Abstracts Service registry numbers.—Continued [Final list compiled according to the National Institute of Standards and Technology (NIST) Web site (http://webbook.nist.gov/chemistry/); NIST Standard Reference Database No. -

Q-1-3-16 Jurado Dictaminador
:, CODIGO B l /3~ ll~LI ~03 UNIVERSIDAD NACIONAL DE SAN AGUSTÍN DE AREQUIPA FACULTAD DE AGRONOMÍA ANÁLISIS TÉCNICO-ECONÓMICO DEL CULTIVO DE QUINUA ORGÁNICA (Chenopodium quinoa Willd.) EN EL DISTRITO DE MAÑAZO DEPARTAMENTO DE PUNO Tesis presentada por la Bachiller: JENNY ELIZABETH HINOJOSA VELÁSQUEZ Para optar el titulo profesional de: INGENIERO AGRÓNOMO AREQUIPAP PERÚ 2015 Ubicacion Física- B-1 Q-1-3-16 JURADO DICTAMINADOR lng. María Antonieta Cahuana Parada lng. José Pinto Cáceres Secretaria Integrante EPÍGRAFE El campo del intelectual es por definición la conciencia. Un Intelectual que no comprende lo que pasa en su tiempo y en su país es una contradicción andante, y el que comprendiendo no actúa, tendrá un lugar en la antología de/llanto, no en la historia viva de su tierra". Rodolfo Walsh (Periódico de la CGT, 1968) DEDICATORIA Dedico este trabajo principalmente a Dios y a la Virgen de Chapi quienes iluminan mi vida, mis ideas, mis pensamientos y me brinda energías para una lucha interminable de mis metas, proyectos y éxitos. A mis padres Vicente y Aleja, toda mi gratitud, por ser /os pilares de mi formación, por su amor, comprensión, compañía, y orientación. A mi esposo Jonathan, por apoyarme en todo /os momentos de mi vida y por haber estado al pendiente de mis actividades, por la motivación para la búsqueda de nuevos horizontes, por su empeño incansable y el respaldo constante a este proyecto. AGRADECIMIENTOS En primera instancia agradezco la dedicación que tuvieron mi asesor y jurados, de la Universidad Nacional de San Agustín de Arequipa quienes aportaron su valiosa experiencia investigativa y tiempo en las revisiones de la presente tesis.