Modeling Diverse Communities of Marine Microbes
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Trophic Tapestry of the Sea
COMMENTARY The trophic tapestry of the sea Matthew J. Church1 Department of Oceanography, University of Hawaii, 1000 Pope Road, Honolulu, HI 96822 ceanic ecosystems are the as a powerful proxy for defining micro- largest habitats on Earth, bial lifestyles. and living biomass in these systems is dominated by Location, Location, Location Oplanktonic microorganisms. Microbial To the casual observer ocean ecosys- ecologists have long sought to under- tems may appear relatively homogenous; stand the role of environmental in reality, these environments are ripe selection in shaping the diversity of with habitat heterogeneity. Climate forc- planktonic microorganisms. The oceans ing of ocean currents, wind-driven mix- have proven a dauntingly complex arena ing, stratification, and upwelling all can to conduct such studies. Marine ecosys- contribute to seascape variability (7). tems are immense and remote and Moreover, numerous biotic processes hence chronically undersampled. More- result in patchy resource distributions. over, the enormous diversity of plank- Dead or decaying plankton cells, excre- tonic microorganisms challenges our ment, and spillage of cellular contents most sophisticated computational capa- into seawater by grazing or viral lysis all bilities. Only recently, through creative can form transient ‘‘hot spots’’ of nutri- applications of new technologies have ents and energy substrates against an oceanographers begun to overcome some Fig. 1. Representation of the ocean as a microbial otherwise dilute resource pool (8, 9). habitat; vertical gradients in sunlight are superim- of these hurdles. In this issue of PNAS, Singularly or together these processes Lauro et al. (1) use a comparative ge- posed on depth-dependent changes in inorganic nutrient concentrations (black line). -

'The Paradox of the Plankton'.Pdf
ecological complexity 4 (2007) 26–33 27 4. Additional limiting factors (density dependent effects) . 30 4.1. Behavioural effects . ......................... 30 4.2. Interactions with others . ...................... 30 4.3. Self limitation by toxin-producing phytoplankton . 30 5. Discussions . ................................... 31 Acknowledgements . ............................ 32 References . .................................. 32 1. Introduction recent approach on this topic. Instead of emphasizing any particle class of mechanisms in detail, we try to present in Phytoplankton are the basis of most aquatic food chains. Most brief the importance of all the mechanisms regulating the of the species of phytoplankton are phototrophs. These plankton dynamics and diversity in real world. phototrophic phytoplankton species ‘‘ . reproduce and build up populations in inorganic media containing a source of CO 2, inorganic nitrogen, sulphur and phosphorous compounds and 2. Classification of the proposed mechanisms a considerable number of other elements (Na, K, Mg, Ca, Si, Fe, Mn, B, Cl, Cu, Zn, Mo, Co and V) most of which are required in Because the principle of competitive exclusion says that the small concentrations and not all of which are known to be number of coexisting species in equilibrium cannot exceed the required by all groups’’ (Hutchinson, 1961) . However, in many number of limiting factors, there may be, in principle, two natural waters, only nitrate, phosphate, light and carbon are possible solutions of the plankton paradox: limiting resources regulating phytoplankton growth. The principle of competitive exclusion (Hardin, 1960; Armstrong (i) due to some reasons, the dynamics of real-world plankton and McGehee, 1980) suggests that in homogeneous, well- never approach to the equilibrium; mixed environments, species that compete for the same (ii) there exist some additional limiting factors that regulate resource cannot coexist, and that in such competitions one the overall dynamics. -

Diversity in Different Trophic Levels of the Plankton
Biogeosciences Discuss., 5, 4897–4917, 2008 Biogeosciences www.biogeosciences-discuss.net/5/4897/2008/ Discussions BGD © Author(s) 2008. This work is distributed under 5, 4897–4917, 2008 the Creative Commons Attribution 3.0 License. Biogeosciences Discussions is the access reviewed discussion forum of Biogeosciences Diversity in different trophic levels of the Similar patterns of community plankton organization characterize distinct groups V. Raybaud et al. of different trophic levels in the plankton Title Page of the NW Mediterranean Sea Abstract Introduction V. Raybaud1,2, A. Tunin-Ley1,3, M. E. Ritchie4, and J. R. Dolan1,3 Conclusions References 1UPMC Univ Paris 6, UMR 7093, Laboratoire d’Oceanographie´ de Villefranche, Observatoire Tables Figures Oceanologique´ de Villefranche-sur-Mer, Station Zoologique, B.P. 28, 06230 Villefranche-Sur-Mer, France J I 2CNRS, UMR 7093, Laboratoire d’Oceanographie´ de Villefranche, Observatoire Oceanologique´ de Villefranche-sur-Mer, Station Zoologique, B.P. 28, 06230 J I Villefranche-Sur-Mer, France 3CNRS, UMR 7093, Laboratoire d’Oceanographie´ de Villefranche, Microbial Ecology and Back Close Biogeochemistry, Observatoire Oceanologique´ de Villefranche-sur-Mer, Station Zoologique, B.P. 28, 06230 Villefranche-Sur-Mer, France Full Screen / Esc 4Biology Department, Syracuse University, Syracuse, NY, USA Printer-friendly Version Received: 24 September 2008 – Accepted: 23 October 2008 – Published: 12 December 2008 Correspondence to: J. R. Dolan ([email protected]) Interactive Discussion Published by Copernicus Publications on behalf of the European Geosciences Union. 4897 Abstract BGD Planktonic populations were sampled over a 4 week period in the NW Mediterranean, at a site subject to little vertical advection during the Dynaproc 2 cruise in 2004. -

Sheldon Spectrum and the Plankton Paradox: Two Sides of the Same Coin : a Trait-Based Plankton Size-Spectrum Model
This is a repository copy of Sheldon Spectrum and the Plankton Paradox: Two Sides of the Same Coin : A trait-based plankton size-spectrum model. White Rose Research Online URL for this paper: https://eprints.whiterose.ac.uk/116959/ Version: Accepted Version Article: Cuesta, Jose A., Delius, Gustav Walter orcid.org/0000-0003-4092-8228 and Law, Richard orcid.org/0000-0002-5550-3567 (2017) Sheldon Spectrum and the Plankton Paradox: Two Sides of the Same Coin : A trait-based plankton size-spectrum model. Journal of Mathematical Biology. pp. 67-96. ISSN 1432-1416 https://doi.org/10.1007/s00285-017-1132-7 Reuse This article is distributed under the terms of the Creative Commons Attribution (CC BY) licence. This licence allows you to distribute, remix, tweak, and build upon the work, even commercially, as long as you credit the authors for the original work. More information and the full terms of the licence here: https://creativecommons.org/licenses/ Takedown If you consider content in White Rose Research Online to be in breach of UK law, please notify us by emailing [email protected] including the URL of the record and the reason for the withdrawal request. [email protected] https://eprints.whiterose.ac.uk/ Journal of Mathematical Biology manuscript No. (will be inserted by the editor) Sheldon Spectrum and the Plankton Paradox: Two Sides of the Same Coin A trait-based plankton size-spectrum model Jose´ A. Cuesta · Gustav W. Delius · Richard Law Received: date / Accepted: date Abstract The Sheldon spectrum describes a remarkable regularity in aquatic ecosys- tems: the biomass density as a function of logarithmic body mass is approximately constant over many orders of magnitude. -

Phytoplankton Sampling in Quantitative Baseline and Monitoring Programs
W&M ScholarWorks Reports 2-1978 Phytoplankton Sampling in Quantitative Baseline and Monitoring Programs Paul E. Stofan Virginia Institute of Marine Science George C. Grant Virginia Institute of Marine Science Follow this and additional works at: https://scholarworks.wm.edu/reports Part of the Marine Biology Commons Recommended Citation Stofan, P. E., & Grant, G. C. (1978) Phytoplankton Sampling in Quantitative Baseline and Monitoring Programs. Special Scientific Report: no. 85. Virginia Institute of Marine Science, College of William and Mary. http://dx.doi.org/doi:10.21220/m2-r64t-f518 This Report is brought to you for free and open access by W&M ScholarWorks. It has been accepted for inclusion in Reports by an authorized administrator of W&M ScholarWorks. For more information, please contact [email protected]. EPA-600/3-78-025 February 1978 Ecological Research Series RESEARCH REPORTING SERIES Research reports of the Office of Research and Development. U.S. Environmental Protection Agency, have been grouped into nine series. These nine broad cate gories were established to facilitate further development and application of en vironmental technology. Elimination of traditional grouping was consciously planned to foster technology transfer and a maximum interface in related fields. The nine series are: 1. Environmental Health Effects Research 2. Environmental Protection Technology 3. Ecological Research 4. Environmental Monitoring 5. Socioeconomic Environmental Studies 6. Scientific and Technical Assessment Reports (STAR) 7. Interagency Energy-Environment Research and Development 8. "Special" Reports 9. Miscellaneous Reports This report has been assigned to thE~ ECOLOGICAL RESEARCH series. This series describes research on the effects of pollution on humans, plant and animal spe cies, and materials. -

Universal Patterns of Matter and Energy Fluxes in Land and Ocean Ecosystems A
bioRxiv preprint doi: https://doi.org/10.1101/2020.04.13.039156; this version posted April 13, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. Universal patterns of matter and energy fluxes in land and ocean ecosystems A. V. Nefiodov Petersburg Nuclear Physics Institute named by B. P. Konstantinov of NRC «Kurchatov Institute», Gatchina, St. Petersburg, 188300, Russia E-mail: [email protected] Abstract. In work [1], the fundamental relationships for the fluxes of matter and energy in terrestrial ecosystems were obtained. Taking into account the universal characteristics of biota, these relationships permitted an estimate to be made of the vertical thickness of the live biomass layer for autotrophs and heterotrophs. The distribution of consumption of biota production as dependent on the body size of heterotrophs was also investigated. For large animals (vertebrates), the energy consumption in sustainable ecosystems was estimated to be of the order of one percent of primary production. In this comment, it is shown that the results of work [1] also hold true for ocean ecosystems and thus are universal for life as a whole. This is of paramount importance for human life on Earth. Keywords: biotic regulation, energy flux, photosynthesis, metabolism, productivity, autotrophs, heterotrophs, plankton On the system of units and dimensions of physical quantities Since the density of living organisms practically coincides with the density of liquid water 1 t/m3, live body mass m and characteristic body volume l 3 are 3 related as m l . -

Searching Explanations of Nature in the Mirror World of Math
Table of Contents Searching Explanations of Nature in the Mirror World of Math........................................................................0 ABSTRACT...................................................................................................................................................0 INTRODUCTION.........................................................................................................................................0 FLIPPING LAKES........................................................................................................................................1 THE TROPHIC CASCADE..........................................................................................................................6 CHAOS IN PLANKTON DYNAMICS......................................................................................................12 SO, WHAT DO MODELS REALLY TELL US?.......................................................................................13 THEORY AS A BRIDGE BETWEEN DISCIPLINES..............................................................................15 RESPONSES TO THIS ARTICLE.............................................................................................................16 Acknowledgments:.......................................................................................................................................16 LITERATURE CITED................................................................................................................................17 -

Fractal Hypothesis of the Pelagic Microbial Ecosystem—Can Simple Ecological Principles Lead to Self-Similar Complexity in the Pelagic Microbial Food Web?
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Frontiers - Publisher Connector HYPOTHESIS AND THEORY published: 01 December 2015 doi: 10.3389/fmicb.2015.01357 Fractal Hypothesis of the Pelagic Microbial Ecosystem—Can Simple Ecological Principles Lead to Self-Similar Complexity in the Pelagic Microbial Food Web? Selina Våge * and T. Frede Thingstad Marine Microbial Ecology Group, Department of Biology, University of Bergen and Hjort Centre for Marine Ecosystem Dynamics, Bergen, Norway Trophic interactions are highly complex and modern sequencing techniques reveal enormous biodiversity across multiple scales in marine microbial communities. Within the chemically and physically relatively homogeneous pelagic environment, this calls for an explanation beyond spatial and temporal heterogeneity. Based on observations of simple parasite-host and predator-prey interactions occurring at different trophic Edited by: Jakob Pernthaler, levels and levels of phylogenetic resolution, we present a theoretical perspective on this University of Zurich, Switzerland enormous biodiversity, discussing in particular self-similar aspects of pelagic microbial Reviewed by: food web organization. Fractal methods have been used to describe a variety of Ryan J. Newton, natural phenomena, with studies of habitat structures being an application in ecology. University of Wisconsin -Milwaukee, USA In contrast to mathematical fractals where pattern generating rules are readily known, Gianluca Corno, however, identifying mechanisms that lead to natural fractals is not straight-forward. National Research Council of Italy - Institute of Ecosystem Study, Italy Here we put forward the hypothesis that trophic interactions between pelagic microbes *Correspondence: may be organized in a fractal-like manner, with the emergent network resembling the Selina Våge structure of the Sierpinski triangle. -
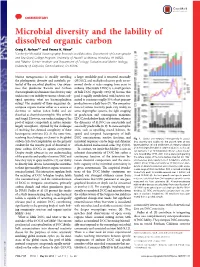
Microbial Diversity and the Lability of Dissolved Organic Carbon Craig E
COMMENTARY Microbial diversity and the lability of dissolved organic carbon Craig E. Nelsona,1 and Emma K. Wearb aCenter for Microbial Oceanography: Research and Education, Department of Oceanography and Sea Grant College Program, University of HawaiʻiatManoa, Honolulu, HI 96822; and bMarine Science Institute and Department of Ecology, Evolution and Marine Biology, University of California, Santa Barbara, CA 93106 Marine metagenomics is steadily unveiling a larger semilabile pool is removed seasonally the phylogenetic diversity and metabolic po- (SLDOC), and multiple refractory pools are re- tential of the microbial plankton. Our aware- moved slowly at scales ranging from years to ness that planktonic Bacteria and Archaea millenia. Observable LDOC is a small portion (bacterioplankton) dominate this diversity only of bulk DOC (typically <2%) (2) because this underscores our inability to answer a basic ecol- pool is rapidly metabolized, with bacteria esti- ogical question: what are bacterioplankton mated to consume roughly 50% of net primary eating? The majority of these organisms de- production on a daily basis (7). The concentra- compose organic matter either as a source of tions of various reactivity pools vary widely; in electrons or carbon (often both) and are some oligotrophic systems, the tight coupling classified as chemoheterotrophic (like animals of production and consumption maintains and fungi). However, our understanding of the LDOC stocks below limits of detection, whereas pool of organic compounds in nature remains thedynamicsofSLDOCaremeasurableand largely amorphous, stymied by the challenges seasonally predictable (8, 9). In more eutrophic of resolving the chemical complexity of these areas, such as upwelling coastal habitats, the heterogenous mixtures (1).Atthesametime, spatial and temporal heterogeneity of bulk resolving these linkages is relevant to the global DOC concentrations, reactive fractions, and Fig. -

Hutchinson's Heritage: the Diversity-Disturbance Relationship In
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by OceanRep Hydrobiologia 249: 1-7, 1993. J. Padisctk, C.S. Reynolds & U. Sommer (eds), Intermediate Disturbance Hypothesis in Phytoplankton Ecology. 1 0 1993 Kluwev Academic Publishers. Printed in Belgium. Hutchinson’s heritage: the diversity-disturbance relationship in phytoplankton U. Sommer’, J. Padisak2, C. S. Reynolds3 & P. Juhasz-Nagy4 1Institut fir Biologie und Chemie des Meeres, Universitat Oldenburg, PO@ 2503, D-2900 Oldenburg, Germany; 2Botanical Department of the Hungarian Natural History Museum/Balaton Limnological Institute of the Hungarian Academy of Sciences, H-8237 Tihany, Hungary; 3Freshwater Biological Association, NERC Institute of Freshwater Ecology, Windermere Laboratory, Ambleside, LA22 OLP, UK; 4Department of Plant Taxonomy and Ecology, Eiitviis Lorand University, HI083 Budapest, Ludovika te’r 2, Hungary Key words: the paradox of plankton, Intermediate Disturbance Hypothesis, diversity, general ecology, community changes Abstract This paper introduces a collection of contributions presented at the 8th Workshop of the International Association of Phytoplankton Taxonomy and Ecology. It compares the substance of with what to limnologists is the more familiar ‘paradox of the plankton’ posed by G. E. Hutchinson. The utility of Connell‘s Intermediate Disturbance Hypothesis in plankton ecology is, potentially, more instructive but inherent difficulties in relating response to stimulus have to be overcome. A copy of the brief distributed to contributors before the workshop is appended. Preamble ilimnetic phytoplankton coexist in a well mixed environment and compete for a very small num- All modern students of limnology will have ben- ber of common limiting resources (light and a few efited from the influence of the late Professor nutritional elements). -

Plankton Dynamics:The Influence of Light; Nutrients and Diversity
PLANKTON DYNAMICS: THE INFLUENCE OF LIGHT, NUTRIENTS AND DIVERSITY Dissertation zur Erlangung des Doktorgrades der Naturwissenschaften Dr. rer. nat. der Fakultät für Biologie der Ludwig-Maximilians-Universität München vorgelegt von Maren Striebel Zur Beurteilung eingereicht am 31. Juli 2008 Tag der mündlichen Prüfung: 07. November 2008 Gutachter: Erstgutachter: PD Dr. Herwig Stibor Zweitgutachter: Prof. Dr. Susanne Foitzik CONTENTS CONTENTS 3 Summary………………..............……………….……………………………………………...... 1. Introduction…………………………………………………………...….…………….............. 7 Phytoplankton, photosynthesis and photosynthetic pigments…...…….…................. 7 Phytoplankton, light and nutrients.……...……………………….................................. 9 The light-nutrient hypothesis………...………………………………………………….... 11 Phytoplankton biodiversity, resource use and productivity.………….……………….. 12 Mobility in phytoplankton species: Advantages and costs…..…………………..……. 14 Estimation of phytoplankton growth and mortality.….....................................….….... 14 2. Papers…………………………………..…………………………………………...………..… 18 2.1 Paper 1: Colorful niches link biodiversity to carbon dynamics in pelagic ecosystems …………….……………......................................................................... 18 2.2 Paper 2: The coupling of biodiversity and productivity in phytoplankton communities: Consequences for biomass stoichiometry……..…………….…….….. 41 2.3 Paper 3 : Light induced changes of plankton growth and stoichiometry: Experiments with natural phytoplankton communities………………………………. -

Lumpy Species Coexistence Arises Robustly in Fluctuating Resource Environments
Lumpy species coexistence arises robustly in fluctuating resource environments Athanasia Sakavaraa,b, George Tsirtsisc, Daniel L. Roelked,e, Rebecca Mancyb, and Sofie Spatharisb,f,1 aDepartment of the Environment, University of the Aegean, Mytilene 81100, Greece; bInstitute of Biodiversity, Animal Health & Comparative Medicine, University of Glasgow, Glasgow G12 8QQ, Scotland, United Kingdom; cDepartment of Marine Sciences, University of the Aegean, Mytilene 81100, Greece; dDepartment of Wildlife and Fisheries Sciences, Texas A&M University, College Station, TX 77843-2258; eDepartment of Oceanography, Texas A&M University, College Station, TX 77843-2258; and fSchool of Life Sciences, University of Glasgow, Glasgow G12 8QQ, Scotland, United Kingdom Edited by Nils Chr. Stenseth, University of Oslo, Oslo, Norway, and approved November 21, 2017 (received for review April 12, 2017) The effect of life-history traits on resource competition outcomes abilities (species A is a superior competitor for resource 2 and is well understood in the context of a constant resource supply. species B for resource 1; Fig. 1A), coexistence is stable when the However, almost all natural systems are subject to fluctuations of resources are supplied within the region of coexistence (dark resources driven by cyclical processes such as seasonality and tidal shading, Fig. 1A) (5). However, when resources fluctuate, the hydrology. To understand community composition, it is therefore resource ratio may fall outside the region of coexistence, at least imperative to study the impact of resource fluctuations on interspe- for some portion of the time. In such scenarios, predicting which cies competition. We adapted a well-established resource-competition species will persist in the long term becomes more challenging.