Edinburgh Research Explorer
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Download Download
Biodiversity Observations http://bo.adu.org.za An electronic journal published by the Animal Demography Unit at the University of Cape Town The scope of Biodiversity Observations consists of papers describing observations about biodiversity in general, including animals, plants, algae and fungi. This includes observations of behaviour, breeding and flowering patterns, distributions and range extensions, foraging, food, movement, measurements, habitat and colouration/plumage variations. Biotic interactions such as pollination, fruit dispersal, herbivory and predation fall within the scope, as well as the use of indigenous and exotic species by humans. Observations of naturalised plants and animals will also be considered. Biodiversity Observations will also publish a variety of other interesting or relevant biodiversity material: reports of projects and conferences, annotated checklists for a site or region, specialist bibliographies, book reviews and any other appropriate material. Further details and guidelines to authors are on this website. Paper Editor: Les G. Underhill OVERVIEW OF THE DISCOVERY OF THE WEAVERS H. Dieter Oschadleus Recommended citation format: Oschadleus HD 2016. Overview of the discovery of the weavers. Biodiversity Observations 7. 92: 1–15. URL: http://bo.adu.org.za/content.php?id=285 Published online: 13 December 2016 – ISSN 2219-0341 – Biodiversity Observations 7.92: 1–15 1 TAXONOMY Currently, 117 living species of weavers in the Ploceidae family are recognised. Hoyo et al. OVERVIEW OF THE DISCOVERY OF THE WEAVERS (2010) listed 116 species but Safford & Hawkins (2013) split the Aldabra Fody Foudia H. Dieter Oschadleus aldabrana from the Red- headed Fody Foudia Animal Demography Unit, Department of Biological Sciences, eminentissima. Dickinson & University of Cape Town, Rondebosch, 7701 South Africa Christidis (2014) also listed 117 species. -

Bontebok Birds
Birds recorded in the Bontebok National Park 8 Little Grebe 446 European Roller 55 White-breasted Cormorant 451 African Hoopoe 58 Reed Cormorant 465 Acacia Pied Barbet 60 African Darter 469 Red-fronted Tinkerbird * 62 Grey Heron 474 Greater Honeyguide 63 Black-headed Heron 476 Lesser Honeyguide 65 Purple Heron 480 Ground Woodpecker 66 Great Egret 486 Cardinal Woodpecker 68 Yellow-billed Egret 488 Olive Woodpecker 71 Cattle Egret 494 Rufous-naped Lark * 81 Hamerkop 495 Cape Clapper Lark 83 White Stork n/a Agulhas Longbilled Lark 84 Black Stork 502 Karoo Lark 91 African Sacred Ibis 504 Red Lark * 94 Hadeda Ibis 506 Spike-heeled Lark 95 African Spoonbill 507 Red-capped Lark 102 Egyptian Goose 512 Thick-billed Lark 103 South African Shelduck 518 Barn Swallow 104 Yellow-billed Duck 520 White-throated Swallow 105 African Black Duck 523 Pearl-breasted Swallow 106 Cape Teal 526 Greater Striped Swallow 108 Red-billed Teal 529 Rock Martin 112 Cape Shoveler 530 Common House-Martin 113 Southern Pochard 533 Brown-throated Martin 116 Spur-winged Goose 534 Banded Martin 118 Secretarybird 536 Black Sawwing 122 Cape Vulture 541 Fork-tailed Drongo 126 Black (Yellow-billed) Kite 547 Cape Crow 127 Black-shouldered Kite 548 Pied Crow 131 Verreauxs' Eagle 550 White-necked Raven 136 Booted Eagle 551 Grey Tit 140 Martial Eagle 557 Cape Penduline-Tit 148 African Fish-Eagle 566 Cape Bulbul 149 Steppe Buzzard 572 Sombre Greenbul 152 Jackal Buzzard 577 Olive Thrush 155 Rufous-chested Sparrowhawk 582 Sentinel Rock-Thrush 158 Black Sparrowhawk 587 Capped Wheatear -

Biodiversity Observations
Biodiversity Observations http://bo.adu.org.za An electronic journal published by the Animal Demography Unit at the University of Cape Town The scope of Biodiversity Observations consists of papers describing observations about biodiversity in general, including animals, plants, algae and fungi. This includes observations of behaviour, breeding and flowering patterns, distributions and range extensions, foraging, food, movement, measurements, habitat and colouration/plumage variations. Biotic interactions such as pollination, fruit dispersal, herbivory and predation fall within the scope, as well as the use of indigenous and exotic species by humans. Observations of naturalised plants and animals will also be considered. Biodiversity Observations will also publish a variety of other interesting or relevant biodiversity material: reports of projects and conferences, annotated checklists for a site or region, specialist bibliographies, book reviews and any other appropriate material. Further details and guidelines to authors are on this website. Paper Editor: Doug Harebottle BIRDS AND ANIMALS USING WEAVERS NESTS H. Dieter Oschadleus Recommended citation format: Oschadleus HD 2017. Birds and animals using weavers nests. Biodiversity Observations, Vol 8.28: 1-17 URL: http://bo.adu.org.za/content.php?id=323 Published online: 20 June 2017 Appendix added: 26 June 2017 – ISSN 2219-0341 – Biodiversity Observations 8.28: 1-17 1 PHOWN (PHOtos of Weaver Nests) Methods BIRDS AND ANIMALS USING WEAVERS NESTS The PHOWN database began in mid July 2010 and the data for this analysis was extracted up to 2 February 2017, providing 6.5 years of H. Dieter Oschadleus data collection. Records with Nest Use were marked so that they could be easily extracted. -

South Africa Mega Birding III 5Th to 27Th October 2019 (23 Days) Trip Report
South Africa Mega Birding III 5th to 27th October 2019 (23 days) Trip Report The near-endemic Gorgeous Bushshrike by Daniel Keith Danckwerts Tour leader: Daniel Keith Danckwerts Trip Report – RBT South Africa – Mega Birding III 2019 2 Tour Summary South Africa supports the highest number of endemic species of any African country and is therefore of obvious appeal to birders. This South Africa mega tour covered virtually the entire country in little over a month – amounting to an estimated 10 000km – and targeted every single endemic and near-endemic species! We were successful in finding virtually all of the targets and some of our highlights included a pair of mythical Hottentot Buttonquails, the critically endangered Rudd’s Lark, both Cape, and Drakensburg Rockjumpers, Orange-breasted Sunbird, Pink-throated Twinspot, Southern Tchagra, the scarce Knysna Woodpecker, both Northern and Southern Black Korhaans, and Bush Blackcap. We additionally enjoyed better-than-ever sightings of the tricky Barratt’s Warbler, aptly named Gorgeous Bushshrike, Crested Guineafowl, and Eastern Nicator to just name a few. Any trip to South Africa would be incomplete without mammals and our tally of 60 species included such difficult animals as the Aardvark, Aardwolf, Southern African Hedgehog, Bat-eared Fox, Smith’s Red Rock Hare and both Sable and Roan Antelopes. This really was a trip like no other! ____________________________________________________________________________________ Tour in Detail Our first full day of the tour began with a short walk through the gardens of our quaint guesthouse in Johannesburg. Here we enjoyed sightings of the delightful Red-headed Finch, small numbers of Southern Red Bishops including several males that were busy moulting into their summer breeding plumage, the near-endemic Karoo Thrush, Cape White-eye, Grey-headed Gull, Hadada Ibis, Southern Masked Weaver, Speckled Mousebird, African Palm Swift and the Laughing, Ring-necked and Red-eyed Doves. -

Geographic Variation in Breeding Seasonality and Primary Moult Parameters in Cape Weavers, Southern Masked Weavers and Southern Red Bishops in South Africa
Chapter 5 Geographic variation in breeding seasonality and primary moult parameters in Cape Weavers, Southern Masked Weavers and Southern Red Bishops in South Africa 97 98 Geographic variation in breeding seasonality and primary moult parameters in Cape Weavers, Southern Masked Weavers and Southern Red Bishops in South Africa Abstract The Southern Masked Weaver Ploceus velatus expanded its range into the south- western part of the Western Cape, and here it has advanced its peak breeding and moult onset by one month relative to other areas, but is still a month behind that of the Cape Weaver P. capensis. Peak breeding in the Southern Red Bishop Euplectes orix is the same as that of the two Ploceus species in KwaZulu-Natal, and retarded by one or two months in the other regions. Southern Masked Weavers use trees and reeds equally early in different parts of South Africa. Variation in start and end of breeding depends largely on rainfall. In the three ploceids, primary moult started in the same month that the last eggs are laid. For the Cape Weaver moult started progressively later along the coast from the Western Cape to KwaZulu-Natal, and duration of primary moult varied from 3.3 months to 4.1 months. In Southern Masked Weavers moult started between 27 December and 22 March, while duration of moult was 2.2 to 2.8 months. In Southern Red Bishops moult started early in the Western Cape (13 December), and much later in the other regions. Duration of moult varied from 2.4 to 3.1 months. -

Common Birds of Namibia and Botswana 1 Josh Engel
Common Birds of Namibia and Botswana 1 Josh Engel Photos: Josh Engel, [[email protected]] Integrative Research Center, Field Museum of Natural History and Tropical Birding Tours [www.tropicalbirding.com] Produced by: Tyana Wachter, R. Foster and J. Philipp, with the support of Connie Keller and the Mellon Foundation. © Science and Education, The Field Museum, Chicago, IL 60605 USA. [[email protected]] [fieldguides.fieldmuseum.org/guides] Rapid Color Guide #584 version 1 01/2015 1 Struthio camelus 2 Pelecanus onocrotalus 3 Phalacocorax capensis 4 Microcarbo coronatus STRUTHIONIDAE PELECANIDAE PHALACROCORACIDAE PHALACROCORACIDAE Ostrich Great white pelican Cape cormorant Crowned cormorant 5 Anhinga rufa 6 Ardea cinerea 7 Ardea goliath 8 Ardea pupurea ANIHINGIDAE ARDEIDAE ARDEIDAE ARDEIDAE African darter Grey heron Goliath heron Purple heron 9 Butorides striata 10 Scopus umbretta 11 Mycteria ibis 12 Leptoptilos crumentiferus ARDEIDAE SCOPIDAE CICONIIDAE CICONIIDAE Striated heron Hamerkop (nest) Yellow-billed stork Marabou stork 13 Bostrychia hagedash 14 Phoenicopterus roseus & P. minor 15 Phoenicopterus minor 16 Aviceda cuculoides THRESKIORNITHIDAE PHOENICOPTERIDAE PHOENICOPTERIDAE ACCIPITRIDAE Hadada ibis Greater and Lesser Flamingos Lesser Flamingo African cuckoo hawk Common Birds of Namibia and Botswana 2 Josh Engel Photos: Josh Engel, [[email protected]] Integrative Research Center, Field Museum of Natural History and Tropical Birding Tours [www.tropicalbirding.com] Produced by: Tyana Wachter, R. Foster and J. Philipp, -

Full Magazine November 2018
Promerops: Magazine of the Cape Bird Club Issue No. 312, November 2018 PO BOX 2113 CLAREINCH 7740 Website: www.capebirdclub.org.za e-Mail: [email protected] CAPE BIRD CLUB COMMITTEE MEMBERS AND OTHER OFFICE BEARERS Hon. President Peter Steyn 021 685 3049 [email protected] COMMITTEE MEMBERS Chairman Priscilla Beeton 084 803 9987 [email protected] Vice-Chairman Johan Schlebusch 021 685 7418 [email protected] Treasurer Joy Fish 021 685 7418 [email protected] Secretary Barbara Jones 021 705 9948 [email protected] Membership Linda Merrett 021 685 7368 [email protected] Conservation Dave Whitelaw 021 671 3714 [email protected] e-Newsletter Cheryl Leslie 082 924 7035 [email protected] Projects & Support Gillian Barnes 021 782 5429 [email protected] Youth & Education Melodie Hertslet 082 255 2062 [email protected] OTHER OFFICE BEARERS Information Sylvia Ledgard ————— [email protected] Book Sales Gavin & Anne Greig 021 794 7791 [email protected] Camps, Local Gillian Barnes 021 782 5429 [email protected] Courses Johan Schlebusch 021 685 7418 [email protected] Media Liaison Linda Johnston 021 712 3707 [email protected] Meetings, Outings Priscilla Beeton 084 803 9987 [email protected] Promerops Fiona Jones 021 671 2594 [email protected] Shop Paul & Cathy Jenkins 021 689 3513 [email protected] Sightings Simon Fogarty 021 701 6303 [email protected] Webmaster Gavin Lawson 021 705 5224 [email protected] THE CAPE BIRD CLUB IS AFFILIATED TO BIRDLIFE SOUTH AFRICA Please send your news, articles, travel reports, observations, Editing & Production: and comments to Fiona Jones at [email protected]. -

Species List
Kenya: Species List Mammals Specie Seen Location Aberdare mole shrew African bush elephant African long-fingered bat African savanna hare African striped weasel Asian house shrew Audacious mole-rat Banded mongoose Bat-eared fox Black-backed jackal Bunyoro rabbit Bushbuck Bushveld horseshoe bat Common duiker Coypu Cushioned gerbil Dik-dik Duiker East African wild dog Echo (elephant) Embi mole-rat Four-toed elephant shrew Grant's gazelle Grant's zebra Greater kudu Grévy's zebra Ground pangolin Hartebeest Heart-nosed bat Hirola Hylomyscus endorobae Kenya coast galago Kenyan African mole-rat Kirk's dik-dik Klipspringer Lesser kudu Maneless zebra Masai giraffe Mianzini mole-rat Mount Kenya mole shrew Naivasha mole-rat Nandi Bear Nyiro shrew Percival's trident bat Peters's flat-headed bat Rainey's shrew Reticulated giraffe Roan antelope Rothschild's giraffe Sable antelope Serval Side-striped jackal Single-striped grass mouse Smoky white-toothed shrew Southern giant pouched rat Steenbok Storey's African mole-rat Tachyoryctes rex Taita shrew Tana River red colobus Thomson's gazelle Tree pangolin Ultimate shrew Waterbuck Yellow-backed duiker Birds Specie Seen Location Common Quail Harlequin Quail Blue Quail Stone Partridge Helmeted Guineafowl Crested Guineafowl Vulturine Guineafowl Fulvous Whistling-Duck White-faced Whistling-Duck White-backed Duck Maccoa Duck Egyptian Goose Ruddy Shelduck Spur-winged Goose Comb Duck African Pygmy-Goose Cape Teal Gadwall Eurasian Wigeon African Black Duck Mallard Yellow-billed Duck Northern Shoveler Red-billed -

SOUTH AFRICA: LAND of the ZULU 26Th October – 5Th November 2015
Tropical Birding Trip Report South Africa: October/November 2015 A Tropical Birding CUSTOM tour SOUTH AFRICA: LAND OF THE ZULU th th 26 October – 5 November 2015 Drakensberg Siskin is a small, attractive, saffron-dusted endemic that is quite common on our day trip up the Sani Pass Tour Leader: Lisle Gwynn All photos in this report were taken by Lisle Gwynn. Species pictured are highlighted RED. 1 www.tropicalbirding.com +1-409-515-0514 [email protected] Page Tropical Birding Trip Report South Africa: October/November 2015 INTRODUCTION The beauty of Tropical Birding custom tours is that people with limited time but who still want to experience somewhere as mind-blowing and birdy as South Africa can explore the parts of the country that interest them most, in a short time frame. South Africa is, without doubt, one of the most diverse countries on the planet. Nowhere else can you go from seeing Wandering Albatross and penguins to seeing Leopards and Elephants in a matter of hours, and with countless world-class national parks and reserves the options were endless when it came to planning an itinerary. Winding its way through the lush, leafy, dry, dusty, wet and swampy oxymoronic province of KwaZulu-Natal (herein known as KZN), this short tour followed much the same route as the extension of our South Africa set departure tour, albeit in reverse, with an additional focus on seeing birds at the very edge of their range in semi-Karoo and dry semi-Kalahari habitats to add maximum diversity. KwaZulu-Natal is an oft-underrated birding route within South Africa, featuring a wide range of habitats and an astonishing diversity of birds. -

Biodiversity Observations
Biodiversity Observations http://bo.adu.org.za An electronic journal published by the Animal Demography Unit at the University of Cape Town The scope of Biodiversity Observations consists of papers describing observations about biodiversity in general, including animals, plants, algae and fungi. This includes observations of behaviour, breeding and flowering patterns, distributions and range extensions, foraging, food, movement, measurements, habitat and colouration/plumage variations. Biotic interactions such as pollination, fruit dispersal, herbivory and predation fall within the scope, as well as the use of indigenous and exotic species by humans. Observations of naturalised plants and animals will also be considered. Biodiversity Observations will also publish a variety of other interesting or relevant biodiversity material: reports of projects and conferences, annotated checklists for a site or region, specialist bibliographies, book reviews and any other appropriate material. Further details and guidelines to authors are on this website. Lead Editor: Arnold van der Westhuizen – Paper Editor: Les G Underhill LONGEVITY RECORDS OF SOUTHERN AFRICAN WEAVERS H. Dieter Oschadleus Recommended citation format: Oschadleus HD 2016. Longevity records of southern African weavers. Biodiversity Observations 7.59: 1–18. URL: http://bo.adu.org.za/content.php?id=252 Published online: 31 August 2016 – ISSN 2219-0341 – Biodiversity Observations 7.59: 1–18 1 AFRING NEWS PAPER Methods LONGEVITY RECORDS OF SOUTHERN AFRICAN The SAFRING database was searched for longevity records for each weaver species in southern Africa. These records were checked WEAVERS against the original records as far as possible. The number of ringed, recaptured and recovered weavers was extracted on 7 August 2016 H. -

Northern South Africa Trip Report Private Tour
NORTHERN SOUTH AFRICA TRIP REPORT PRIVATE TOUR 31 OCTOBER – 5 NOVEMBER 2017 By Dylan Vasapolli The rare Taita Falcon (Falco fasciinucha) showed well. www.birdingecotours.com [email protected] 2 | TRIP REPORT Northern South Africa November 2017 Overview This short tour was an extension to northern South Africa for a private client, following our set- departure South Africa tours to the Western Cape and Subtropical South Africa. The primary goal was to target species occurring in northern South Africa that are absent/uncommon elsewhere in the country and/or any species missed on the set-departure South Africa tours. This short tour began and ended in Johannesburg and saw us transiting northwards first to the rich thornveld of the Zaagkuilsdrift Road, followed by the montane forests of the Magoebaskloof hills before visiting the moist grasslands and broad-leaved woodlands found in north-eastern Gauteng. Day 1, October 31. Johannesburg Following the conclusion of our set-departure Subtropical South Africa tour earlier in the day I met up with James, who would be joining this post-tour extension, in the early evening. We headed for dinner and, following dinner, drove south of Johannesburg to search for African Grass Owl. We arrived on site and began working the region. We enjoyed excellent views of a number of Marsh Owls, along with Spotted Thick-knee, but drew a blank on the grass owl for most of our time. But at our last stop we had success and saw a single African Grass Owl come in, although it remained distant and did leave us wanting more. -
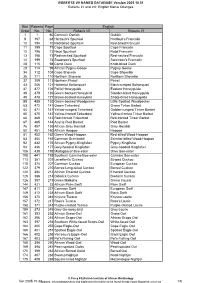
English Name Changes Bird Roberts Page Order No. No. Roberts V
ROBERTS VII NAMES DATABASE Version 2005.10.31 Roberts VI and VII: English Name Changes Bird Roberts Page English Order No. No. Roberts VII Roberts VI 1 1 60 Common Ostrich Ostrich 9 197 68 Hartlaub's Spurfowl Hartlaub's Francolin 10 194 70 Red-billed Spurfowl Red-billed Francolin 11 195 71 Cape Spurfowl Cape Francolin 12 196 72 Natal Spurfowl Natal Francolin 13 198 73 Red-necked Spurfowl Red-necked Francolin 14 199 74 Swainson's Spurfowl Swainson's Francolin 28 115 98 Comb Duck Knob-billed Duck 29 114 99 African Pygmy-Goose Pygmy Goose 34 112 108 Cape Shoveler Cape Shoveller 35 111 110 Northern Shoveler Northern Shoveller 37 109 113 Northern Pintail Pintail 43 206 121 Hottentot Buttonquail Black-rumped Buttonquail 47 477 126 Pallid Honeyguide Eastern Honeyguide 48 479 126 Green-backed Honeybird Slender-billed Honeyguide 49 478 127 Brown-backed Honeybird Sharp-billed Honeyguide 55 485 133 Green-backed Woodpecker Little Spotted Woodpecker 63 472 141 Green Tinkerbird Green Tinker Barbet 64 471 141 Yellow-rumped Tinkerbird Golden-rumped Tinker Barbet 65 470 142 Yellow-fronted Tinkerbird Yellow-fronted Tinker Barbet 66 469 143 Red-fronted Tinkerbird Red-fronted Tinker Barbet 67 465 144 Acacia Pied Barbet Pied Barbet 76 457 155 African Grey Hornbill Grey Hornbill 80 451 160 African Hoopoe Hoopoe 81 452 162 Green Wood-Hoopoe Red-billed Wood-Hoopoe 83 454 165 Common Scimitarbill Scimitar-billed Wood-Hoopoe 92 432 176 African Pygmy-Kingfisher Pygmy Kingfisher 93 436 177 Grey-headed Kingfisher Grey-hooded Kingfisher 106 439 192 Madagascar Bee-eater