Distribution Patterns and Habitat Use of the Smooth Hammerhead Shark (Sphyrna Zygaena) in the Atlantic Ocean
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Scalloped Hammerhead Shark (Sphyrna Lewini) 2014-2019 Bibliography
Scalloped hammerhead shark (Sphyrna lewini) 2014-2019 Bibliography Trevor Riley, Head of Public Services, NOAA Central Library NCRL subject guide 2019-09 https://doi.org/10.25923/79kf-v153 September 2019 U.S. Department of Commerce National Oceanic and Atmospheric Administration Office of Oceanic and Atmospheric Research NOAA Central Library – Silver Spring, Maryland Table of Contents Background & Scope ................................................................................................................................. 3 Sources Reviewed ..................................................................................................................................... 3 Section I: Biology and Life History ............................................................................................................. 4 Section II: Genetics .................................................................................................................................. 17 Section III: Population Abundance .......................................................................................................... 20 Section IV: Threats .................................................................................................................................. 28 2 Background & Scope Scalloped hammerhead sharks are moderately large sharks with a global distribution. The most distinguishing characteristic of this shark is its "hammer-shaped" head. They are threatened by commercial fishing, mainly for the shark fin trade. Two distinct -

NPOA Sharks Booklet.Indd
National Plan of Action for the Conservation and Management of Sharks (NPOA-Sharks) November 2013 South Africa Department of Agriculture, Forestry and Fisheries Private Bag X2, Rogge Bay, 8012 Tel: 021 402 3911 Fax: +27 21 402 3364 www.daff.gov.za Design and Layout: FNP Communications and Gerald van Tonder Photographs courtesy of: Department of Agriculture, Forestry and Fisheries (DAFF), Craig Smith, Charlene da Silva, Rob Tarr Foreword South Africa’s Exclusive Economic Zone is endowed with a rich variety of marine living South Africa is signatory to the Code of Conduct for Responsible Fisheries – voluntarily agreed to by members of the United Nations Food and Agriculture Organisation (FAO) – and, as such, is committed to the development and implementation of National Plans of Action (NPOAs) as adopted by the twenty-third session of the FAO Committee on Fisheries in February 1999 and endorsed by the FAO Council in June 1999. Seabirds – aimed at reducing incidental catch and promoting the conservation of seabirds Fisheries and now regularly conducts Ecological Risk Assessments for all the commercial practices. Acknowledging the importance of maintaining a healthy marine ecosystem and the possibility of major detrimental effects due to the disappearance of large predators, South from the list of harvestable species. In accordance with international recommendations, South Africa subsequently banned the landing of a number of susceptible shark species, including oceanic whitetip, silky, thresher and hammerhead sharks. improves monitoring efforts for foreign vessels discharging shark products in its ports. To ensure long-term sustainability of valuable, but biologically limited, shark resources The NPOA-Sharks presented here formalises and streamlines ongoing efforts to improve conservation and management of sharks caught in South African waters. -

Reproductive Biology of the Bonnethead (Sphyrna Tiburo) from the Southeastern U.S
University of North Florida UNF Digital Commons UNF Graduate Theses and Dissertations Student Scholarship 2014 Reproductive Biology of the Bonnethead (Sphyrna tiburo) from the Southeastern U.S. Atlantic Coast Melissa I. Gonzalez De Acevedo University of North Florida, [email protected] Follow this and additional works at: https://digitalcommons.unf.edu/etd Part of the Biology Commons, and the Ecology and Evolutionary Biology Commons Suggested Citation Gonzalez De Acevedo, Melissa I., "Reproductive Biology of the Bonnethead (Sphyrna tiburo) from the Southeastern U.S. Atlantic Coast" (2014). UNF Graduate Theses and Dissertations. 534. https://digitalcommons.unf.edu/etd/534 This Master's Thesis is brought to you for free and open access by the Student Scholarship at UNF Digital Commons. It has been accepted for inclusion in UNF Graduate Theses and Dissertations by an authorized administrator of UNF Digital Commons. For more information, please contact Digital Projects. © 2014 All Rights Reserved REPRODUCTIVE BIOLOGY OF THE BONNETHEAD (SPHYRNA TIBURO) FROM THE SOUTHEASTERN U.S. ATLANTIC COAST by Melissa Gonzalez De Acevedo A thesis submitted to the Department of Biology in partial fulfillment of the requirements for the degree of Masters of Science in Biology UNIVERSITY OF NORTH FLORIDA COLLEGE OF ARTS AND SCIENCES December 2014 Unpublished work, © Melissa Gonzalez De Acevedo CERTIFICATE OF APPROVAL The thesis “Reproductive biology of the bonnethead (Sphyrna tiburo) from the southeastern U.S. Atlantic coast” submitted by Melissa Gonzalez De Acevedo Approved by the thesis committee: Date Dr. Jim Gelsleichter Committee Chair Dr. Carolyn Belcher Dr. Eric Johnson Accepted for the Department of Biology: Dr. Cliff Ross Assistant Chair Accepted for the College of Arts and Sciences: Dr. -

The Bonnethead (Sphyrna Tiburo) and the Scalloped Hammerhead (Sphyrna Lewini) Sarah L
© 2017. Published by The Company of Biologists Ltd | Journal of Experimental Biology (2017) 220, 3336-3343 doi:10.1242/jeb.157941 RESEARCH ARTICLE Regional variation in undulatory kinematics of two hammerhead species: the bonnethead (Sphyrna tiburo) and the scalloped hammerhead (Sphyrna lewini) Sarah L. Hoffmann1,*, Steven M. Warren2 and Marianne E. Porter1 ABSTRACT blochii), possesses a cephalofoil that is proportionally the largest Hammerhead sharks (Sphyrnidae) exhibit a large amount of and measures up to 50% of their total body length (Lim et al., 2010). morphological variation within the family, making them the focus of In comparison, the bonnethead shark (Sphyrna tiburo) is the most many studies. The size of the laterally expanded head, or cephalofoil, is recently derived species and their cephalofoil width is 18% of total inversely correlated with pectoral fin area. The inverse relationship body length. Generally, as cephalofoil width increases among between cephalofoil and pectoral fin size in this family suggests that they species, pectoral fin area decreases (Thomson and Simanek, 1977). might serve a complementary role in lift generation. The cephalofoil is Previous studies on hammerhead sharks have focused primarily on also hypothesized to increase olfaction, electroreception and vision; cephalofoil morphology and its effects on hydrodynamics and however, little is known about how morphological variation impacts post- sensory efficiency; however, little is known about the morphology cranial swimming kinematics. Previous studies demonstrate that the and function of the post-cranial body. The significant bonnethead and scalloped hammerhead have significantly different yaw morphological variation and the close phylogenetic relationship amplitude, and we hypothesized that these species utilize varied among hammerheads make them an ideal study system to examine frequency and amplitude of undulation along the body. -
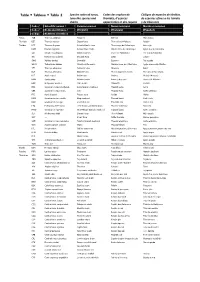
Table Tableau Tabla 2
Table Tableau Tabla 2 Species codes of tunas, Codes des espèces de Códigos de especies de túnidos, tuna‐like species and thonidés, d’espèces de especies afines a los túnidos sharks apparentées et des requins y de tiburones Code / Scientific names / Common names Noms communs Nombres comunes Code / Noms sientifiques / (English) (Français) (Español) Código Nombres científicos Tunas ALB Thunnus alalunga Albacore Germon Atún blanco Thonidés BET Thunnus obesus Bigeye tuna Thon obèse(=Patudo) Patudo Túnidos BFT Thunnus thynnus Atlantic bluefin tuna Thon rouge de l’atlantique Atún rojo BUM Makaira nigricans Atlantic blue marlin Makaire bleu de l'Atlantique Aguja azul del Atlántico SAI Istiophorus albicans Atlantic sailfish Voilier de l'Atlantique Pez vela del Atlántico SKJ Katsuwonus pelamis Skipjack tuna Listao Listado SWO Xiphias gladius Swordfish Espadon Pez espada WHM Tetrapturus albidus Atlantic white marlin Makaire blanc de l'Atlantique Aguja blanca del Atlántico YFT Thunnus albacares Yellowfin tuna Albacore Rabil BLF Thunnus atlanticus Blackfin tuna Thon à nageoires noires Atún des aletas negras BLT Auxis rochei Bullet tuna Bonitou Melva(=Melvera) BON Sarda sarda Atlantic bonito Bonite à dos rayé Bonito del Atlántico BOP Orcynopsis unicolor Plain bonito Palomette Tasarte BRS Scomberomorus brasiliensis Serra Spanish mackerel Thazard serra Serra CER Scomberomorus regalis Cero Thazard franc Carite chinigua FRI Auxis thazard Frigate tuna Auxide Melva KGM Scomberomorus cavalla King mackerel Thazard barré Carite lucio KGX Scomberomorus spp -

Sphyrna Tudes (Smalleye Hammerhead Shark)
UWI The Online Guide to the Animals of Trinidad and Tobago Ecology Sphyrna tudes (Smalleye Hammerhead Shark) Family: Sphyrnidae (Hammerhead Sharks) Order: Carcharhiniformes (Ground Sharks) Class: Chondrichthyes (Cartilaginous Fish) Fig. 1. Smalleye hammerhead shark, Sphyrna tudes. [http://otlibrary.com/wp-content/gallery/golden-hammerhead-shark/hammerhead-jeff-pierce-lo-res.jpg, downloaded 7 January 2015] TRAITS. One of the smaller species of hammerhead (Family Sphyrnidae), the smalleye hammerhead shark can reach a length of 1.5m and weigh around 9 kg. They have a streamlined, slender body as opposed to their bulkier relatives (Martin, 1999). Adult males may range from 110-130cm total length while adult females have an average length of 120-145cm. Newborns are usually about 30cm at birth. Common to all hammerhead sharks is the mallet/hammer shaped cephalofoil, which has a width of 28-32% of body length, and is wide and long. Newborns typically have a longer cephalofoil, which is more curved and with a smaller indent than that of adults (Castro, 1989). These hammerheads have considerably smaller eyes compared to that of other hammerheads, hence its name, and have tri-layered eyelids for protection. However the most distinctive trait being their golden coloration (Fig. 1), which gives the species other names like the golden hammerhead or the curry shark (Gallagher, 2010). Their colour may range from UWI The Online Guide to the Animals of Trinidad and Tobago Ecology bright gold to orange-yellow; however these colours only appear at the juvenile stage, usually when a length of 45cm is reached, and fade at sexual maturity (Castro, 1989). -

The Social Lives of Hammerheads. Authors: De Maddalena, Alessandro; Buttigieg, Alexander Publication: World and I Online Date: Jun 1, 2006
The social lives of hammerheads. Authors: De Maddalena, Alessandro; Buttigieg, Alexander Publication: World and I Online Date: Jun 1, 2006 Hammerhead sharks form the family of Sphyrnidae, that includes eight species: the winghead shark (Eusphyra blochii), scalloped bonnethead (Sphyrna corona), scalloped hammerhead (S. lewini), scoophead shark (S. media), great hammerhead (S. mokarran), bonnethead shark (S. tiburo), golden hammerhead (S. tudes) and the smooth hammerhead (S. zygaena). In the hammerhead shark, the front part of the head is flattened dorsoventrally and laterally increased to form what we call the "cephalofoil," or two wide flattened expansions that constitute the characteristic shape of a hammer. These two expansions are made out of muscular and connective tissues supported by a cartilaginous skeleton that is an integrated part of the skull. The round shaped eyes are placed very wide apart at the two lateral extremities of these expansions. Also placed at the front margin of these lateral expansions are the nostrils. The "hammer" or cephalofoil of the hammerheads unfolds into a series of functions correlated to the movement and the predation of these sharks. It has been observed that hammerhead sharks posses a very highly developed brain, and perhaps this could be the reason for the ability to lead the social life particular to these animals. Without a doubt, the most surprising aspect of socialization in hammerhead sharks is that of the immensely huge gatherings that different species form in numerous geographical areas. These gatherings could be formed from resident populations in that area or can be composed of individuals that migrate in masses. Huge gatherings of scalloped hammerhead sharks have been observed at the Sea of Cortez, near the Galapagos Island, near Cocos Island (Costa Rica), Malpelo (Columbia), and San Salvador (Bahamas), Hawaii, the Red Sea, Natal (South Africa), in Australia, the China Sea and near Cabilao Island (Philippines). -

(Orcinus Orca) and Hammerhead Sharks (Sphyrna Sp.) in Galápagos Waters
LAJAM 5(1): 69-71, June 2006 ISSN 1676-7497 INTERACTION BETWEEN KILLER WHALES (ORCINUS ORCA) AND HAMMERHEAD SHARKS (SPHYRNA SP.) IN GALÁPAGOS WATERS LUCA SONNINO SORISIO1, ALESSANDRO DE MADDALENA2 AND INGRID N. VISSER3,4 ABSTRACT: A possible predatory interaction between killer whales (Orcinus orca) and hammerhead sharks (Sphyrna sp.) was observed during April 1991 near Punta Cormorant, Galápagos Islands. Three killer whales were observed in close proximity to a freshly dead female hammerhead. One of the killer whales (approximately 6m in length) was observed motionless in a vertical position above the shark carcass and later was seen chasing an approximately 40cm hammerhead, supposedly a pup born prematurely from the dead shark. The sharks are thought to have been scalloped hammerheads (S. lewini). RESÚMEN: Una posible interacción predatoria entre orcas (Orcinus orca) y peces martillo (Sphyrna sp.) fue observada en Abril de 1991 cerca de Punta Cormorán, Islas Galápagos. Tres orcas fueron vistas muy próximas a una hembra de pez martillo recién muerta. Una de las orcas (de unos 6m de longitud), fue observada inmóvil en posición vertical sobre la carcasa del tiburón y después fue vista persiguiendo a un pez martillo de unos 40cm, supuestamente una cría nacida prematuramente de la hembra muerta. Se piensa que los tiburones pudieran ser cornudas negras (S. lewini). KEYWORDS: killer whale; Orcinus orca; hammerhead shark; Sphyrna; predation. Observations of killer whales (Orcinus orca) feeding on, the infrequent sightings and even rarer number of or attacking sharks are relatively infrequent (Table 1). observations of killer whale predation off the Galápagos Reports of killer whales off the Galápagos Islands are Islands, we report here on a possible predatory not common (e.g., Day, 1994; Merlen, 1999; Smith and interaction between killer whales and hammerhead Whitehead, 1999) as during a 27-year period of record sharks (Sphyrna sp.). -

Great Hammerhead Shark (Sphyrna Mokarran) UNDER the U.S
PETITION TO LIST THE Great Hammerhead Shark (Sphyrna mokarran) UNDER THE U.S. ENDANGERED SPECIES ACT Photo: Gary J. Wood (creative commons license) Petition Submitted to the U.S. Secretary of Commerce, Acting Through the National Oceanic and Atmospheric Administration and the National Marine Fisheries Service Petitioner: WildEarth Guardians 1536 Wynkoop Street, Suite 301 Denver, CO 80202 (303) 573-4898 December 18, 2012 INTRODUCTION WildEarth Guardians hereby formally petitions the Secretary of Commerce (Secretary), acting through the National Marine Fisheries Service (NMFS), an agency within the National Oceanic and Atmospheric Administration (NOAA), to list the great hammerhead shark (Sphyrna mokarran) as “threatened” or “endangered” under the U.S. Endangered Species Act (ESA) (16 U.S.C. §§ 1531-1544). We request that NMFS list the species throughout its range; however, in the alternative, if NMFS finds that there are Distinct Population Segments (DPS) of great hammerhead sharks, we would request that those be listed under the ESA. Additionally, we request that NMFS designate critical habitat for the species in U.S. waters or areas of the high seas that are essential to the species’ survival and recovery. The great hammerhead shark is the largest of all hammerhead sharks and is found in warm temperate and tropical waters around the world. Great hammerhead populations are in severe decline; the International Union for Conservation of Nature (IUCN) lists great hammerhead sharks as “endangered” on the IUCN Red List. IUCN Red List 2010a, Exhibit 1 at 1. The species faces at least five major threats. The first is the present and threatened destruction of great hammerhead habitat by pollution and anthropogenic climate change. -

Species Composition of the Largest Shark Fin Retail-Market in Mainland
www.nature.com/scientificreports OPEN Species composition of the largest shark fn retail‑market in mainland China Diego Cardeñosa1,2*, Andrew T. Fields1, Elizabeth A. Babcock3, Stanley K. H. Shea4, Kevin A. Feldheim5 & Demian D. Chapman6 Species‑specifc monitoring through large shark fn market surveys has been a valuable data source to estimate global catches and international shark fn trade dynamics. Hong Kong and Guangzhou, mainland China, are the largest shark fn markets and consumption centers in the world. We used molecular identifcation protocols on randomly collected processed fn trimmings (n = 2000) and non‑ parametric species estimators to investigate the species composition of the Guangzhou retail market and compare the species diversity between the Guangzhou and Hong Kong shark fn retail markets. Species diversity was similar between both trade hubs with a small subset of species dominating the composition. The blue shark (Prionace glauca) was the most common species overall followed by the CITES‑listed silky shark (Carcharhinus falciformis), scalloped hammerhead shark (Sphyrna lewini), smooth hammerhead shark (S. zygaena) and shortfn mako shark (Isurus oxyrinchus). Our results support previous indications of high connectivity between the shark fn markets of Hong Kong and mainland China and suggest that systematic studies of other fn trade hubs within Mainland China and stronger law‑enforcement protocols and capacity building are needed. Many shark populations have declined in the last four decades, mainly due to overexploitation to supply the demand for their fns in Asia and meat in many other countries 1–4. Mainland China was historically the world’s second largest importer of shark fns and foremost consumer of shark fn soup, yet very little is known about the species composition of shark fns in this trade hub2. -
Elasmobranchs (Sharks and Rays): a Review of Status, Distribution and Interaction with Fisheries in the Southwest Indian Ocean
See discussions, stats, and author profiles for this publication at: http://www.researchgate.net/publication/277329893 Elasmobranchs (sharks and rays): a review of status, distribution and interaction with fisheries in the Southwest Indian Ocean CHAPTER · JANUARY 2015 READS 81 2 AUTHORS, INCLUDING: Jeremy J Kiszka Florida International University 52 PUBLICATIONS 389 CITATIONS SEE PROFILE Available from: Jeremy J Kiszka Retrieved on: 16 October 2015 OFFSHORE FISHERIES OF THE SOUTHWEST INDIAN OCEAN: their status and the impact on vulnerable species OCEANOGRAPHIC RESEARCH INSTITUTE Special Publication No. 10 Rudy van der Elst and Bernadine Everett (editors) The Investigational Report series of the Oceanographic Research Institute presents the detailed results of marine biological research. Reports have appeared at irregular intervals since 1961. All manuscripts are submitted for peer review. The Special Publication series of the Oceanographic Research Institute reports on expeditions, surveys and workshops, or provides bibliographic and technical information. The series appears at irregular intervals. The Bulletin series of the South African Association for Marine Biological Research is of general interest and reviews the research and curatorial activities of the Oceanographic Research Institute, uShaka Sea World and the Sea World Education Centre. It is published annually. These series are available in exchange for relevant publications of other scientific institutions anywhere in the world. All correspondence in this regard should be directed to: The Librarian Oceanographic Research Institute PO Box 10712 Marine Parade 4056 Durban, South Africa OFFSHORE FISHERIES OF THE SOUTHWEST INDIAN OCEAN: their status and the impact on vulnerable species Rudy van der Elst and Bernadine Everett (editors) South African Association for Marine Biological Research Oceanographic Research Institute Special Publication No. -

Sphyrna Zygaena
Published Date: 1 March 2019 Smooth Hammerhead, Sphyrna zygaena Report Card Sustainable assessment IUCN Red List IUCN Red List Australian Near Threatened Global Vulnerable Assessment Assessment Simpfendorfer, C., Gaibor, N., Soldo, A., Heupel, M.R., Smith, W.D., Assessors Stevens, J.D. & Vooren, C.M. In Australia, severe declines in NSW but stable in the remainder of Report Card Remarks Australian range Summary The Smooth Hammerhead is one of the larger Source: Clinton Duffy/Fishbase. License: CC BY hammerhead sharks and is distributed worldwide. The Attribution-NonCommercial fins are highly valued and the species is caught with a wide variety of gears in both coastal and oceanic fisheries, as bycatch and a target. There is limited data on the species’ life history, though it is presumably at least as biologically sensitive as the Scalloped Hammerhead. Species-specific data are often not available to assess population trends as hammerhead species are mostly grouped but hammerheads show dramatic population declines of >99% in some areas. On the basis of these estimated reductions in several locations and suspected declines in other areas, the species is assessed as globally Endangered (IUCN). In Australia, there are conflicting trends in the population between the east (declining) and west coasts (stable or increasing). Data from the New South Wales shark control program shows an approximate 85% decline in general hammerhead catch rates, while the species-specific catch data from the southwest coast gillnet fishery suggests the population is stable or increasing. When the population trend is weighted according to the relative size of NSW and the remainder of the Australian distribution, the total population decline in Australia over three generations is estimated to be 20%.