The Gross and Microscopic Structure of the Hamster Eye with Special
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Eyelid Conjunctival Tumors
EYELID &CONJUNCTIVAL TUMORS PHOTOGRAPHIC ATLAS Dr. Olivier Galatoire Dr. Christine Levy-Gabriel Dr. Mathieu Zmuda EYELID & CONJUNCTIVAL TUMORS 4 EYELID & CONJUNCTIVAL TUMORS Dear readers, All rights of translation, adaptation, or reproduction by any means are reserved in all countries. The reproduction or representation, in whole or in part and by any means, of any of the pages published in the present book without the prior written consent of the publisher, is prohibited and illegal and would constitute an infringement. Only reproductions strictly reserved for the private use of the copier and not intended for collective use, and short analyses and quotations justified by the illustrative or scientific nature of the work in which they are incorporated, are authorized (Law of March 11, 1957 art. 40 and 41 and Criminal Code art. 425). EYELID & CONJUNCTIVAL TUMORS EYELID & CONJUNCTIVAL TUMORS 5 6 EYELID & CONJUNCTIVAL TUMORS Foreword Dr. Serge Morax I am honored to introduce this Photographic Atlas of palpebral and conjunctival tumors,which is the culmination of the close collaboration between Drs. Olivier Galatoire and Mathieu Zmuda of the A. de Rothschild Ophthalmological Foundation and Dr. Christine Levy-Gabriel of the Curie Institute. The subject is now of unquestionable importance and evidently of great interest to Ophthalmologists, whether they are orbital- palpebral specialists or not. Indeed, errors or delays in the diagnosis of tumor pathologies are relatively common and the consequences can be serious in the case of malignant tumors, especially carcinomas. Swift diagnosis and anatomopathological confirmation will lead to a treatment, discussed in multidisciplinary team meetings, ranging from surgery to radiotherapy. -
Lid and Lash Conditions
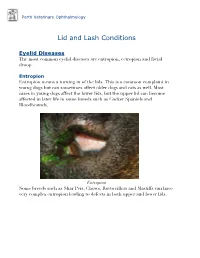
Perth Veterinary Ophthalmology Lid and Lash Conditions Eyelid Diseases The most common eyelid diseases are entropion, ectropion and facial droop. Entropion Entropion means a turning in of the lids. This is a common complaint in young dogs but can sometimes affect older dogs and cats as well. Most cases in young dogs affect the lower lids, but the upper lid can become affected in later life in some breeds such as Cocker Spaniels and Bloodhounds. Entropion Some breeds such as Shar Peis, Chows, Rottweillers and Mastiffs can have very complex entropion leading to defects in both upper and lower lids. A Shar Pei with severe upper and lower lid entropion Entropion is painful and can be potentially blinding. The rolling in of the lid leads to hair coming into contact with the cornea, leading to pain, ulceration and scarring (which can affect vision). In severe cases this can even lead to perforation of the eye. There are many causes of entropion. It can be primary or secondary to other problems affecting the lids (such as ectopic cilia, distichiasis etc. - see below). Some possible causes include the lid being too long, the lid being too tight, instability of the lateral canthus (outer cornea of the eyelids), misdirection of the lateral canthal tendon, brachycephalic anatomy (big eyes and short nose - e.g. Pekingese, Pugs, Shih Tsus, Persian cats etc.), diamond eye defects, loose or too much skin, facial droop etc. Often these cases are referred to a veterinary ophthalmologist for proper assessment and treatment to provide the best outcome. Entropion requires surgical correction. -

The Management of Congenital Malpositions of Eyelids, Eyes and Orbits
Eye (\988) 2, 207-219 The Management of Congenital Malpositions of Eyelids, Eyes and Orbits S. MORAX AND T. HURBLl Paris Summary Congenital malformations of the eye and its adnexa which are multiple and varied can affect the whole eyeball or any part of it, as well as the orbit, eyelids, lacrimal ducts, extra-ocular muscles and conjunctiva. A classification of these malformations is presented together with the general principles of treatment, age of operating and surgical tactics. The authors give some examples of the anatomo-clinical forms, eyelid malpositions such as entropion, ectropion, ptosis, levator eyelid retraction, medial canthus malposition, congenital eyelid colobomas, and congenital orbital abnormalities (Craniofacial stenosis, orbi tal plagiocephalies, hypertelorism, anophthalmos, microphthalmos and cryptophthalmos) . Congenital malformations of the eye and its as echography, CT-scan and NMR, enzymatic adnexa are multiple and varied. They can work-up or genetic studies (Table I). affect the whole eyeball or any part of it, as Surgical treatment when feasible will well as the orbit, eyelids, lacrimal ducts extra encounter numerous problems; age will play a ocular muscles and conjunctiva. role, choice of a surgical protocol directly From the anatomical point of view, the fol related to the existing complaints, and coop lowing can be considered. eration between several surgical teams Position abnormalities (malpositions) of (ophthalmologic, plastic, cranio-maxillo-fac one or more elements and formation abnor ial and neurosurgical), the ideal being to treat malities (malformations) of the same organs. Some of these abnormalities are limited to Table I The manag ement of cong enital rna/positions one organ and can be subjected to a relatively of eyelid s, eyes and orbits simple and well recognised surgical treat Ocular Findings: ment. -

Endoscopic Supraorbital Eyebrow Approach for the Surgical Treatment of Extraaxial and Intraaxial Tumors
See the corresponding editorial in this issue (E21). Neurosurg Focus 37 (4):E20, 2014 ©AANS, 2014 Endoscopic supraorbital eyebrow approach for the surgical treatment of extraaxial and intraaxial tumors ROBERTO GAZZERI, M.D.,1,2 YUYA NISHIYAMA, M.D., PH.D.,1,3 And CHARLES TEO, M.D.1 1Centre for Minimally Invasive Neurosurgery, Prince of Wales Private Hospital, Sydney, Australia; 2Department of Neurosurgery, San Giovanni Addolorata Hospital, Rome, Italy; and 3Department of Neurosurgery, Fujita Health University School of Medicine, Toyoake, Japan Object. The supraorbital eyebrow approach is a minimally invasive technique that offers wide access to the anterior skull base region and parasellar area through a subfrontal corridor. The use of neuroendoscopy allows one to extend the approach further to the pituitary fossa, the anterior third ventricle, the interpeduncular cistern, the an- terior and medial temporal lobe, and the middle fossa. The supraorbital approach involves a limited skin incision, with minimal soft-tissue dissection and a small craniotomy, thus carrying relatively low approach-related morbidity. Methods. All consecutive patients who underwent the endoscopic supraorbital eyebrow approach were retro- spectively analyzed for lesion location, pathology, length of stay, complications, and cosmetic results. Results. During a 56-month period, 97 patients (mean age 58.5 years) underwent an endoscopic eyebrow ap- proach to resect extra- and intraaxial brain lesions. The most common pathologies treated were meningiomas (n = 41); craniopharyngiomas (n = 22); dermoid tumors (n = 7); metastases (n = 4); gliomas (n = 3); and other miscel- laneous frontal, parasellar, and midbrain (n = 23) lesions. The median length of postoperative hospital stay was 2.7 days (range 1–8 days). -

New Theory on Facial Beauty: Ideal Dimensions in the Face and Its Application to Your Practice by Dr
New Theory on Facial Beauty: Ideal Dimensions in the Face And its application to your practice By Dr. Philip Young Aesthetic Facial Plastic Surgery 2015 Bellevue, Washington American Brazilian Aesthetic Meeting • Hello my presentation is on studying some further elements of a new theory on facial beauty called the Circles of Prominence. • Specifically we are going to be studying some key dimensions in the face that I think could possibly help your practice. • I’m from Bellevue Washington Home of Bill Gates, Microsoft and Starbucks. Beauty In my opinion Beauty is the most important trait that we have and it is the one trait that can have the most dramatic impact in our lives. Obviously finding the answer for Beauty is essential in our industry. The answers have alluded us: the magic number of Phi, cephalometrics, the neo classical canons by Leonardo Da Vinci, the averageness theory, etc. have all come short in finding what makes a face beautiful. • The Circles of Prominence is a theory that I discovered in 2003-2005 and was published in the Archives of Facial Plastic Surgery in 2006 and Received the Sir Harold Delf Gillies Award from the American Academy of Facial Plastic Surgery. The Circles of Prominence • Original published Archives FPS 2006 • Based on the idea that there is an ideal • Everything on the face has an ideal as well • Because we spend so much time looking at the iris • All dimensions of the face are related to the width of the iris • Obviously with a better definition of beauty our results in plastic surgery can be improved • The circles of prominence is based on the belief that there is an ideal. -

A Pictorial Anatomy of the Human Eye/Anophthalmic Socket: a Review for Ocularists
A Pictorial Anatomy of the Human Eye/Anophthalmic Socket: A Review for Ocularists ABSTRACT: Knowledge of human eye anatomy is obviously impor- tant to ocularists. This paper describes, with pictorial emphasis, the anatomy of the eye that ocularists generally encounter: the anophthalmic eye/socket. The author continues the discussion from a previous article: Anatomy of the Anterior Eye for Ocularists, published in 2004 in the Journal of Ophthalmic Prosthetics.1 Michael O. Hughes INTRODUCTION AND RATIONALE B.C.O. Artificial Eye Clinic of Washington, D.C. Understanding the basic anatomy of the human eye is a requirement for all Vienna, Virginia health care providers, but it is even more significant to eye care practition- ers, including ocularists. The type of eye anatomy that ocularists know, how- ever, is more abstract, as the anatomy has been altered from its natural form. Although the companion eye in monocular patients is usually within the normal range of aesthetics and function, the affected side may be distorted. While ocularists rarely work on actual eyeballs (except to cover microph- thalmic and blind, phthisical eyes using scleral cover shells), this knowledge can assist the ocularist in obtaining a naturally appearing prosthesis, and it will be of greater benefit to the patient. An easier exchange among ocularists, surgeons, and patients will result from this knowledge.1, 2, 3 RELATIONSHIPS IN THE NORMAL EYE AND ORBIT The opening between the eyelids is called the palpebral fissure. In the nor- mal eye, characteristic relationships should be recognized by the ocularist to understand the elements to be evaluated in the fellow eye. -

Chapter 18 EYELID and ADNEXAL INJURIES
Eyelid and Adnexal Injuries Chapter 18 EYELID AND ADNEXAL INJURIES KIMBERLY PEELE COCKERHAM, MD* INTRODUCTION HISTORY, EXAMINATION, AND ANCILLARY TESTING SURGICAL MANAGEMENT OF EYELID LACERATIONS Anesthesia Preoperative Preparation Superficial Eyelid Lacerations Eyelid-Margin Involvement Levator Injury Complex Eyelid Injuries Total Avulsion SURGICAL MANAGEMENT OF ADNEXAL INJURIES Eyebrow Injuries Canalicular Injuries Medial Canthal Injuries Lateral Canthal Injuries Lacrimal Sac and Lacrimal Gland Injuries POSTOPERATIVE WOUND CARE SUMMARY *Director, Ophthalmic Plastics, Orbital Disease and Neuro-Ophthalmology, Allegheny General Hospital, 420 East North Avenue, Pittsburgh, Pennsylvania 15212; Assistant Professor, Department of Ophthalmology, Drexel University College of Medicine, Philadelphia, Pennsylvania 19102; formerly, Major, Medical Corps, US Army; Director, Neuro-Ophthalmology, Orbital Disease, and Plastic Reconstruction, Depart- ment of Ophthalmology, Walter Reed Army Medical Center, Washington, DC 291 Ophthalmic Care of the Combat Casualty INTRODUCTION Eyelid lacerations are a common emergency injuries are not considered true emergencies, and room challenge. The primary repair, which should in triage settings, life-threatening and actual sight- be the definitive surgery, is too often performed by threatening injuries (ie, open globes) should take medical students or physicians without ophthalmo- precedence. Eyelid lacerations can be closed up to logical training.1 Whether the setting is a civilian 72 hours after the injury without a great impact on emergency room or a battlefield, ophthalmologists functional or aesthetic outcome. Canalicular injury should be involved as early as possible. Inadequate can also be delayed (up to 48 hours), allowing the repair can lead to ocular irritation, pain, and even all-important primary closure to be performed by loss of the eye in cases of severe eyelid dysfunc- an experienced ophthalmologist. -

Anatomy of the Periorbital Region Review Article Anatomia Da Região Periorbital
RevSurgicalV5N3Inglês_RevistaSurgical&CosmeticDermatol 21/01/14 17:54 Página 245 245 Anatomy of the periorbital region Review article Anatomia da região periorbital Authors: Eliandre Costa Palermo1 ABSTRACT A careful study of the anatomy of the orbit is very important for dermatologists, even for those who do not perform major surgical procedures. This is due to the high complexity of the structures involved in the dermatological procedures performed in this region. A 1 Dermatologist Physician, Lato sensu post- detailed knowledge of facial anatomy is what differentiates a qualified professional— graduate diploma in Dermatologic Surgery from the Faculdade de Medician whether in performing minimally invasive procedures (such as botulinum toxin and der- do ABC - Santo André (SP), Brazil mal fillings) or in conducting excisions of skin lesions—thereby avoiding complications and ensuring the best results, both aesthetically and correctively. The present review article focuses on the anatomy of the orbit and palpebral region and on the important structures related to the execution of dermatological procedures. Keywords: eyelids; anatomy; skin. RESU MO Um estudo cuidadoso da anatomia da órbita é muito importante para os dermatologistas, mesmo para os que não realizam grandes procedimentos cirúrgicos, devido à elevada complexidade de estruturas envolvidas nos procedimentos dermatológicos realizados nesta região. O conhecimento detalhado da anatomia facial é o que diferencia o profissional qualificado, seja na realização de procedimentos mini- mamente invasivos, como toxina botulínica e preenchimentos, seja nas exéreses de lesões dermatoló- Correspondence: Dr. Eliandre Costa Palermo gicas, evitando complicações e assegurando os melhores resultados, tanto estéticos quanto corretivos. Av. São Gualter, 615 Trataremos neste artigo da revisão da anatomia da região órbito-palpebral e das estruturas importan- Cep: 05455 000 Alto de Pinheiros—São tes correlacionadas à realização dos procedimentos dermatológicos. -

Tunica Interna (Nervosa) Retina
Visual system 2 and accessory visual structures Organum visuale et structurae oculi accessoriae David Kachlík Tunica interna (nervosa) Retina • pars caeca – pars iridica – pars ciliaris • ora serrata • pars optica – 10 layers – pigmented part – sensory part Retina – pigmented part • stratum pigmentosum • simple cuboid epithelium on basal lamina = Bruch‘s membrane • cells (pigmentocytus) connected by tight junctions • apical parts contain melanin granules • microvilli separate the outer segments of rods and cones • interphotoreceptor matrix (IRBP) • nutrition of rods and cones, photopigment resynthesis, degradation of membranous discs, barrier „blood-retina“ Retina – sensory part • light-sensitive neurons (transducers) – rods and cones • trasmission neurons (integraters) – bipolar and ganglionic cells • association neurons – horizontal and amacrinne cells • suppoting cells (glia) – radial glial cells (Müller´s cells) Section of eyball layers Rods = Neura bacillifera • rod = bacillum retinae • spherula – synaptic terminal • axonal process • nucleus black-abd-white vision • inner segment – GA, ER, MIT; synthesis of ATP and rhodopsin – myoideum (glycogen) + ellipsoideum (mitochondria) • connecting cilium (cilium connectens) – modified cilium • outer segment (segmentum externum) – membranous discs with photopigment – migrate externally and are released Cones = Neuron coniferum • cone = conus retinae • synaptic pedicle (pes terminalis) • photopigment is iodopsin • outer segment – membranous discs with photopigment • communicate with surroundings • color vision – 3 types of cones – according to wave-length – „blue“ – 420 nm – type S – „green“ – 535 nm – type M – „red“ – 565 nm – type L Retina – Transmission neurons • Bipolar neurons (Neuron bipolare) – rod bipolar neurons (n.b. bacillotopicum) – cone bipolar neurons (n.b. conotopicum) • midget (n.b.c. nanum) – macula lutea (no convergence = 1 : 1 : 1) • diffuse (n.b.c. diffusum) – convergence of signal – contact with retinal ganglion cells • Retinal ganglion cells (N. -

Surgical Microanatomy of the Müller Muscle-Conjunctival Resection
ORIGINAL ARTICLE Surgical Microanatomy of the Mu¨ller Muscle-Conjunctival Resection Ptosis Procedure Marcus M. Marcet, M.D.*†, Pete Setabutr, M.D.‡, Bradley N. Lemke, M.D.§, Megan E. Collins, M.D.*, James C. Fleming, M.D.ʈ, Ralph E. Wesley, M.D.¶, Jayant M. Pinto, M.D.*, and Allen M. Putterman, M.D.‡ Departments of *Surgery and †Pathology, University of Chicago; ‡Department of Ophthalmology, University of Illinois, Chicago, Illinois; §Department of Ophthalmology, University of Wisconsin, Madison, Wisconsin; ʈHamilton Eye Institute, University of Tennessee, Memphis; and ¶Vanderbilt University Eye Institute, Nashville, Tennessee, U.S.A. exact mechanism by which the surgery lifts the eyelid has been Purpose: To assess for alterations in the microscopic anat- unclear.2–4 It has been postulated to be secondary to a number omy that occur as a result of the Mu¨ller muscle-conjunctival of factors, including resection and advancement of Mu¨ller resection (MMCR) ptosis procedure and to better understand muscle (MM) and advancement of the levator aponeurosis.2,5 the mechanisms by which MMCR elevates the eyelid. This study uniquely uses a cadaveric model to charac- Methods: Sixteen orbits from 8 fresh frozen Caucasian terize the microanatomic alterations produced with the MMCR cadaver heads, ranging from 38 to 100 years of age were used. procedure. By visualizing the altered microscopic anatomy, we For each head, MMCR was performed on one side. The contralat- hoped to gain insight into the mechanism of action of the eral, unoperated orbit served as an anatomic control. Each exen- surgery. In seeking a visually integrated depiction of collagen, terated orbital contents and excised MMCR specimen was evalu- muscle, and elastic fibers for the histopathologic evaluation, a ated. -

Acquired Etiologies of Lacrimal System Obstructions
5 Acquired Etiologies of Lacrimal System Obstructions Daniel P. Schaefer Acquired obstructions of the lacrimal excretory outfl ow system will produce the symptoms of epiphora, mucopurulent discharge, pain, dacryocystitis, and even cellulitis, prompting the patient to seek the ophthalmologist for evaluation and treatment. Impaired tear outfl ow may be functional, structural, or both. The causes may be primary – those resulting from infl ammation of unknown causes that lead to occlusive fi brosis—or secondary, resulting from infections, infl amma- tion, trauma, malignancies, toxicity, or mechanical causes. Secondary acquired dacryostenosis and obstruction may result from many causes, both common and obscure. Occasionally, the precise pathogenesis of nasolacrimal duct obstruction will, despite years of investigations, be elusive. To properly evaluate and appropriately treat the patient, the ophthal- mologist must have knowledge and comprehension of the lacrimal anatomy, the lacrimal apparatus, pathophysiology, ocular and nasal relationships, ophthalmic and systemic disease process, as well as the topical and systemic medications that can affect the nasolacrimal duct system. One must be able to assess if the cause is secondary to outfl ow anomalies, hypersecretion or refl ex secretion, pseudoepiphora, eyelid malposition abnormalities, trichiasis, foreign bodies and conjunctival concretions, keratitis, tear fi lm defi ciencies or instability, dry eye syn- dromes, ocular surface abnormalities, irritation or tumors affecting the trigeminal nerve, allergy, medications, or environmental factors. Abnormalities of the lacrimal pump function can result from involu- tional changes, eyelid laxity, facial nerve paralysis, or fl oppy eyelid syndrome, all of which displace the punctum from the lacrimal lake. If the cause is secondary to obstruction of the nasolacrimal duct system, the ophthalmologist must be able to determine where the anomaly is and what the cause is, in order to provide the best treatment possible for the patient. -

The Lacrimal System Terms
The Lacrimal System Lynn E. Lawrence, CPOT, ABOC, COA, OSC Terms • Etiology – the cause of a disease or abnormal condition • Dacryocystitis – inflammation of the lacrimal sac • Epiphora – watering of eyes due to excess secretion of tears or obstruction of the lacrimal passage Tear Film Layers oil aqueous snot What functions does each layer of the tear perform? Lacrimal System: Tear Film Layers LIPID DEFICIENCY ‐ evaporates TEAR DEFICIENCY – fails to hydrate properly oil aqueous snot What functions does each layer of the tear perform? What are functions of tears? Tear Components • Lipid Layer – prevents evaporation • Aqueous Layer ‐ hydration • Mucus Layer – sticks tear to the eye • Other components Lacrimal Apparatus • Sometimes a person cannot produce natural tears they might need punctal plugs to prevent the tears from draining off the eye. • Faucet • Action • Drain Obstructive – vs‐ non‐obstructive Tear Production – Secretory • Lacrimal gland – Reflex tearing – Too much tearing…epiphora • Gland of Krause – Superior fornix • Gland of Wolfring – Superior tarsal plate Two Primary Forms of Dry Eye 800 nm 8,000 nm 100 nm The two primary forms of dry eye are Evaporative Dry Eye, also known as Meibomian Gland Dysfunction or MGD and Aqueous Dry Eye. The majority of dry eye sufferers have MGD. Oil & Water Remember science class? Oil floats. Oil does not mix with water, but rather sits on top of water. Oil is what keeps water from evaporating. Need three volunteers TEST TIME http://optometrytimes.modernmedicine.com/optometrytimes/news/treating‐dry‐eye‐ lipid‐based‐eye‐drops Lipid Secretion: Meibomian Glands Left: Transillumination of eyelid showing meibomian glands Right: Secretion of lipid at lid margin • The lipid layer restricts evaporation to 5‐10% of tear flow – Also helps lubricate Mucin Secretion: Goblet Cells Superficial layer of bulbar conjunctiva.