Possibilities for an Aerial Biosphere in Temperate Sub Neptune-Sized Exoplanet Atmospheres
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Water, Habitability, and Detectability Steve Desch
Water, Habitability, and Detectability Steve Desch PI, “Exoplanetary Ecosystems” NExSS team School of Earth and Space Exploration, Arizona State University with Ariel Anbar, Tessa Fisher, Steven Glaser, Hilairy Hartnett, Stephen Kane, Susanne Neuer, Cayman Unterborn, Sara Walker, Misha Zolotov Astrobiology Science Strategy NAS Committee, Beckmann Center, Irvine, CA (remotely), January 17, 2018 How to look for life on (Earth-like) exoplanets: find oxygen in their atmospheres How Earth-like must an exoplanet be for this to work? Seager et al. (2013) How to look for life on (Earth-like) exoplanets: find oxygen in their atmospheres Oxygen on Earth overwhelmingly produced by photosynthesizing life, which taps Sun’s energy and yields large disequilibrium signature. Caveats: Earth had life for billions of years without O2 in its atmosphere. First photosynthesis to evolve on Earth was anoxygenic. Many ‘false positives’ recognized because O2 has abiotic sources, esp. photolysis (Luger & Barnes 2014; Harman et al. 2015; Meadows 2017). These caveats seem like exceptions to the ‘rule’ that ‘oxygen = life’. How non-Earth-like can an exoplanet be (especially with respect to water content) before oxygen is no longer a biosignature? Part 1: How much water can terrestrial planets form with? Part 2: Are Aqua Planets or Water Worlds habitable? Can we detect life on them? Part 3: How should we look for life on exoplanets? Part 1: How much water can terrestrial planets form with? Theory says: up to hundreds of oceans’ worth of water Trappist-1 system suggests hundreds of oceans, especially around M stars Many (most?) planets may be Aqua Planets or Water Worlds How much water can terrestrial planets form with? Earth- “snow line” Standard Sun distance models of distance accretion suggest abundant water. -

2-6-NASA Exoplanet Archive
The NASA Exoplanet Archive Rachel Akeson NASA Exoplanet Science Ins5tute (NExScI) September 29, 2016 NASA Big Data Task Force Overview: Dat a NASA Exoplanet Archive supports both the exoplanet science community and NASA exoplanet missions (Kepler, K2, TESS, WFIRST) • Data • Confirmed exoplanets from the literature • Over 80,000 planetary and stellar parameter values for 3388 exoplanets • Updated weekly • Kepler stellar proper5es, planet candidate, data valida5on and occurrence rate products • MAST is archive for pixel and light curve data • Addi5onal space (CoRoT) and ground-based transit surveys (~20 million light curves) • Transit spectroscopy data • Auto-updated exoplanet plots and movies Big Data Task Force: Exoplanet Archive Overview: Tool s Example: Kepler 14 Time Series Viewer • Interac5ve tables and ploZng for data • Includes light curve normaliza5on • Periodogram calcula5ons • Searches for periodic signals in archive or user-supplied light curves • Transit predic5ons • Uses value from archive to predict future planet transits for observa5on and mission planning • URL-based queries Aliases • Calcula5on of observable proper5es Planet orbital period • Web-based service to collect follow-up observa5ons of planet candidates for Kepler, K2 and TESS (ExoFOP) Stellar ac5vity • Includes user-supplied data, file and notes Big Data Task Force: Exoplanet Archive Data Challenges and Technical Approach (1) • Challenges with Exoplanet Archive are not currently about data volume but about providing CPU resources and data complexity • CPU challenge -
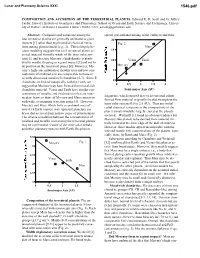
Composition and Accretion of the Terrestrial Planets
Lunar and Planetary Science XXXI 1546.pdf COMPOSITION AND ACCRETION OF THE TERRESTRIAL PLANETS. Edward R. D. Scott and G. Jeffrey Taylor, Hawai’i Institute of Geophysics and Planetology, School of Ocean and Earth Science and Technology, Univer- sity of Hawai’i at Manoa, Honolulu, Hawai’i 96822, USA; [email protected] Abstract: Compositional variations among the spread gravitational mixing of the embryos and their four terrestrial planets are generally attributed to giant 20 impacts [1] rather than to primordial chemical varia- Fig. 2 tions among planetesimals [e.g., 2]. This is largely be- Mars cause modeling suggests that each terrestrial planet ac- 15 creted material from the whole of the inner solar sys- tem [1], and because Mercury’s high density is attrib- 10 Venus Earth uted to mantle stripping in a giant impact [3] and not to its position as the innermost planet [4]. However, Mer- Mercury cury’s high concentration of metallic iron and low con- 5 centration of oxidized iron are comparable to those in recently discovered metal-rich chondrites [5-7]. Since E 0 chondrites are linked isotopically with the Earth, we 0 0.5 1 1.5 2 suggest that Mercury may have formed from metal-rich chondritic material. Venus and Earth have similar con- Semi-major Axis (AU) centrations of metallic and oxidized iron that are inter- fragments, which ensured that each terrestrial planet mediate between those of Mercury and Mars consistent formed from material originally located throughout the with wide, overlapping accretion zones [1]. However, inner solar system (0.5 to 2.5 AU). -

POSSIBLE STRUCTURE MODELS for the TRANSITING SUPER-EARTHS:KEPLER-10B and 11B
43rd Lunar and Planetary Science Conference (2012) 1290.pdf POSSIBLE STRUCTURE MODELS FOR THE TRANSITING SUPER-EARTHS:KEPLER-10b AND 11b. P. Futó1 1 Department of Physical Geography, University of West Hungary, Szombathely, Károlyi Gáspár tér, H- 9700, Hungary ([email protected]) Introduction:Up to january of 2012,10 super- The planet Kepler-11b has a large radius (1.97 R⊕) for Earths have been announced by Kepler-mission [1] its mass (4.3 M⊕),therefore this planet must have a that is designed to detect hundreds of transiting exo- spherical shell that is composed of low-density materi- planets.Kepler was launched on 6th March,2009 and the als.Considering the planet's average density,it must primary purpose of its scientific program is to search have a metallic core with different possible fractional for terrestrial-sized planets in the habitable zone of mass.Accordinghly,I have made a possible structure Solar-like stars.For the case of high number of discov- model for Kepler-11b in which the selected core mass eries we will be able to estimate the frequency of fraction is 32.59% (similarly to that of Earth) and the Earth-sized planets in our galaxy.Results of the Kepler- water ice layer has a relatively great fractional 's measurements show that the small-sized planets are volume.For case of the selected composition, the icy frequent in the spiral galaxies.A catalog of planetary surface sublimated to form a water vapor as the planet candidates,including objects with small-sized candidate moved inward the central star during its migration. -

Progress and Challenges in Using Stable Isotopes to Trace Plant Carbon and Water Relations Across Scales
Biogeosciences, 9, 3083–3111, 2012 www.biogeosciences.net/9/3083/2012/ Biogeosciences doi:10.5194/bg-9-3083-2012 © Author(s) 2012. CC Attribution 3.0 License. Progress and challenges in using stable isotopes to trace plant carbon and water relations across scales C. Werner1,19, H. Schnyder2, M. Cuntz3, C. Keitel4, M. J. Zeeman5, T. E. Dawson6, F.-W. Badeck7, E. Brugnoli8, J. Ghashghaie9, T. E. E. Grams10, Z. E. Kayler11, M. Lakatos12, X. Lee13, C. Maguas´ 14, J. Ogee´ 15, K. G. Rascher1, R. T. W. Siegwolf16, S. Unger1, J. Welker17, L. Wingate18, and A. Gessler11 1Experimental and Systems Ecology, University Bielefeld, 33615 Bielefeld, Germany 2Lehrstuhl fur¨ Grunlandlehre,¨ Technische Universitat¨ Munchen,¨ 85350 Freising-Weihenstephan, Germany 3UFZ – Helmholtz Centre for Environmental Research, Permoserstr. 15, 04318 Leipzig, Germany 4University of Sydney, Faculty of Agriculture, Food and Natural Resources, 1 Central Avenue, Eveleigh, NSW 2015, Australia 5College of Oceanic and Atmospheric Sciences, Oregon State University, 104 COAS Admin Bldg, Corvallis (OR), USA 6Center for Isotope Biogeochemistry, Department of Integrative Biology, University of California, Berkeley, CA 94720, USA 7Potsdam Institute for Climate Impact Research (PIK) PF 60 12 03, 14412 Potsdam, Germany 8CNR – National Research Council of Italy – Institute of Agro-environmental and Forest Biology, via Marconi 2, 05010 Porano (TR), Italy 9Laboratoire d’Ecologie, Systematique´ et Evolution (ESE), CNRS AgroParisTech – UMR8079, Batimentˆ 362, Universite´ de Paris-Sud (XI), 91405 Orsay Cedex, France 10Ecophysiology of Plants, Department of Ecology and Ecosystem Management, Technische Universitat¨ Munchen,¨ Von-Carlowitz-Platz 2, 85354 Freising, Germany 11Institute for Landscape Biogeochemistry Leibniz-Zentrum fur¨ Agrarlandschaftsforschung (ZALF) e.V., Eberswalderstr. -

Planetary Science
Scientific Research National Aeronautics and Space Administration Planetary Science MSFC planetary scientists are active in Peering into the history The centerpiece of the Marshall efforts is research at Marshall, building on a long history of the solar system the Marshall Noble Gas Research Laboratory of scientific support to NASA’s human explo- (MNGRL), a unique facility within NASA. Noble- ration program planning, whether focused Marshall planetary scientists use multiple anal- gas isotopes are a well-established technique on the Moon, asteroids, or Mars. Research ysis techniques to understand the formation, for providing detailed temperature-time histories areas of expertise include planetary sample modification, and age of planetary materials of rocks and meteorites. The MNGRL lab uses analysis, planetary interior modeling, and to learn about their parent planets. Sample Ar-Ar and I-Xe radioactive dating to find the planetary atmosphere observations. Scientists analysis of this type is well-aligned with the formation age of rocks and meteorites, and at Marshall are involved in several ongoing priorities for scientific research and analysis Ar/Kr/Ne cosmic-ray exposure ages to under- planetary science missions, including the Mars in NASA’s Planetary Science Division. Multiple stand when the meteorites were launched from Exploration Rovers, the Cassini mission to future missions are poised to provide new their parent planets. Saturn, and the Gravity Recovery and Interior sample-analysis opportunities. Laboratory (GRAIL) mission to the Moon. Marshall is the center for program manage- ment for the Agency’s Discovery and New Frontiers programs, providing programmatic oversight into a variety of missions to various planetary science destinations throughout the solar system, from MESSENGER’s investiga- tions of Mercury to New Frontiers’ forthcoming examination of the Pluto system. -

Guest Editorial: My Formative Years with the Rasc by Sara Seager I First Learned About Astronomy As a Small Child
INTRODUCTION 9 GUEST EDITORIAL: MY FORMATIVE YEARS WITH THE RASC BY SARA SEAGER I first learned about astronomy as a small child. One of my first memories is looking through a telescope at the Moon. I was completely stunned by what I saw. The Moon— huge and filled with craters—was a world in and of itself. I was with my father, at a star party hosted by the RASC. Later, when I was 10 years old and on my first camping trip, I remember awakening late one night, stepping outside the tent, and looking up. Stars—millions of them it seemed—filled the entire sky and took my breath away. As a teenager in the late 1980s, I joined the RASC Toronto Centre and deter- minedly attended the bimonthly Friday night meetings held in what was then the Planetarium building. Much of the information went over my head, but I can still clearly remember the excitement of the group whenever a visiting professor presented on a novel topic. An RASC member offered a one-semester evening class on astron- omy, and we were able to use the planetarium to learn about the night sky. The RASC events were a highlight of my high school and undergraduate years. While majoring in math and physics at the University of Toronto, I was thrilled to intern for two summers at the David Dunlap Observatory (DDO). I carried out an observing program of variable stars (including Polaris, the North Star), using the 61-cm (24″) and 48-cm (19″) telescopes atop the DDO’s administration building for simultaneous photometry of the target and comparison stars. -

Revisiting the Sulfur-Water Chemical System in the Middle Atmosphere of Venus Wencheng Shao, Xi Zhang, Carver Bierson, Therese Encrenaz
Revisiting the Sulfur-Water Chemical System in the Middle Atmosphere of Venus Wencheng Shao, Xi Zhang, Carver Bierson, Therese Encrenaz To cite this version: Wencheng Shao, Xi Zhang, Carver Bierson, Therese Encrenaz. Revisiting the Sulfur-Water Chemi- cal System in the Middle Atmosphere of Venus. Journal of Geophysical Research. Planets, Wiley- Blackwell, 2020, 125 (8), pp.e06195. 10.1029/2019JE006195. hal-03250477 HAL Id: hal-03250477 https://hal.archives-ouvertes.fr/hal-03250477 Submitted on 11 Jun 2021 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Copyright RESEARCH ARTICLE Revisiting the Sulfur‐Water Chemical System 10.1029/2019JE006195 in the Middle Atmosphere of Venus Key Points: Wencheng D. Shao1 , Xi Zhang1 , Carver J. Bierson1 , and Therese Encrenaz2 • We found that there is no bifurcation behavior in the 1Department of Earth and Planetary Sciences, University of California, Santa Cruz, CA, USA, 2LESIA, Observatoire de sulfur‐water chemical system as previously claimed Paris, PSL University, CNRS, Sorbonne University, University Sorbonne Paris City, Meudon, France • The observed SO2‐H2O anticorrelation can be explained by the sulfur‐water chemistry with Abstract Sulfur‐water chemistry plays an important role in the middle atmosphere of Venus. -

+ New Horizons
Media Contacts NASA Headquarters Policy/Program Management Dwayne Brown New Horizons Nuclear Safety (202) 358-1726 [email protected] The Johns Hopkins University Mission Management Applied Physics Laboratory Spacecraft Operations Michael Buckley (240) 228-7536 or (443) 778-7536 [email protected] Southwest Research Institute Principal Investigator Institution Maria Martinez (210) 522-3305 [email protected] NASA Kennedy Space Center Launch Operations George Diller (321) 867-2468 [email protected] Lockheed Martin Space Systems Launch Vehicle Julie Andrews (321) 853-1567 [email protected] International Launch Services Launch Vehicle Fran Slimmer (571) 633-7462 [email protected] NEW HORIZONS Table of Contents Media Services Information ................................................................................................ 2 Quick Facts .............................................................................................................................. 3 Pluto at a Glance ...................................................................................................................... 5 Why Pluto and the Kuiper Belt? The Science of New Horizons ............................... 7 NASA’s New Frontiers Program ........................................................................................14 The Spacecraft ........................................................................................................................15 Science Payload ...............................................................................................................16 -

Biosignatures Search in Habitable Planets
galaxies Review Biosignatures Search in Habitable Planets Riccardo Claudi 1,* and Eleonora Alei 1,2 1 INAF-Astronomical Observatory of Padova, Vicolo Osservatorio, 5, 35122 Padova, Italy 2 Physics and Astronomy Department, Padova University, 35131 Padova, Italy * Correspondence: [email protected] Received: 2 August 2019; Accepted: 25 September 2019; Published: 29 September 2019 Abstract: The search for life has had a new enthusiastic restart in the last two decades thanks to the large number of new worlds discovered. The about 4100 exoplanets found so far, show a large diversity of planets, from hot giants to rocky planets orbiting small and cold stars. Most of them are very different from those of the Solar System and one of the striking case is that of the super-Earths, rocky planets with masses ranging between 1 and 10 M⊕ with dimensions up to twice those of Earth. In the right environment, these planets could be the cradle of alien life that could modify the chemical composition of their atmospheres. So, the search for life signatures requires as the first step the knowledge of planet atmospheres, the main objective of future exoplanetary space explorations. Indeed, the quest for the determination of the chemical composition of those planetary atmospheres rises also more general interest than that given by the mere directory of the atmospheric compounds. It opens out to the more general speculation on what such detection might tell us about the presence of life on those planets. As, for now, we have only one example of life in the universe, we are bound to study terrestrial organisms to assess possibilities of life on other planets and guide our search for possible extinct or extant life on other planetary bodies. -

Exep Science Plan Appendix (SPA) (This Document)
ExEP Science Plan, Rev A JPL D: 1735632 Release Date: February 15, 2019 Page 1 of 61 Created By: David A. Breda Date Program TDEM System Engineer Exoplanet Exploration Program NASA/Jet Propulsion Laboratory California Institute of Technology Dr. Nick Siegler Date Program Chief Technologist Exoplanet Exploration Program NASA/Jet Propulsion Laboratory California Institute of Technology Concurred By: Dr. Gary Blackwood Date Program Manager Exoplanet Exploration Program NASA/Jet Propulsion Laboratory California Institute of Technology EXOPDr.LANET Douglas Hudgins E XPLORATION PROGRAMDate Program Scientist Exoplanet Exploration Program ScienceScience Plan Mission DirectorateAppendix NASA Headquarters Karl Stapelfeldt, Program Chief Scientist Eric Mamajek, Deputy Program Chief Scientist Exoplanet Exploration Program JPL CL#19-0790 JPL Document No: 1735632 ExEP Science Plan, Rev A JPL D: 1735632 Release Date: February 15, 2019 Page 2 of 61 Approved by: Dr. Gary Blackwood Date Program Manager, Exoplanet Exploration Program Office NASA/Jet Propulsion Laboratory Dr. Douglas Hudgins Date Program Scientist Exoplanet Exploration Program Science Mission Directorate NASA Headquarters Created by: Dr. Karl Stapelfeldt Chief Program Scientist Exoplanet Exploration Program Office NASA/Jet Propulsion Laboratory California Institute of Technology Dr. Eric Mamajek Deputy Program Chief Scientist Exoplanet Exploration Program Office NASA/Jet Propulsion Laboratory California Institute of Technology This research was carried out at the Jet Propulsion Laboratory, California Institute of Technology, under a contract with the National Aeronautics and Space Administration. © 2018 California Institute of Technology. Government sponsorship acknowledged. Exoplanet Exploration Program JPL CL#19-0790 ExEP Science Plan, Rev A JPL D: 1735632 Release Date: February 15, 2019 Page 3 of 61 Table of Contents 1. -

1 the Atmosphere of Pluto As Observed by New Horizons G
The Atmosphere of Pluto as Observed by New Horizons G. Randall Gladstone,1,2* S. Alan Stern,3 Kimberly Ennico,4 Catherine B. Olkin,3 Harold A. Weaver,5 Leslie A. Young,3 Michael E. Summers,6 Darrell F. Strobel,7 David P. Hinson,8 Joshua A. Kammer,3 Alex H. Parker,3 Andrew J. Steffl,3 Ivan R. Linscott,9 Joel Wm. Parker,3 Andrew F. Cheng,5 David C. Slater,1† Maarten H. Versteeg,1 Thomas K. Greathouse,1 Kurt D. Retherford,1,2 Henry Throop,7 Nathaniel J. Cunningham,10 William W. Woods,9 Kelsi N. Singer,3 Constantine C. C. Tsang,3 Rebecca Schindhelm,3 Carey M. Lisse,5 Michael L. Wong,11 Yuk L. Yung,11 Xun Zhu,5 Werner Curdt,12 Panayotis Lavvas,13 Eliot F. Young,3 G. Leonard Tyler,9 and the New Horizons Science Team 1Southwest Research Institute, San Antonio, TX 78238, USA 2University of Texas at San Antonio, San Antonio, TX 78249, USA 3Southwest Research Institute, Boulder, CO 80302, USA 4National Aeronautics and Space Administration, Ames Research Center, Space Science Division, Moffett Field, CA 94035, USA 5The Johns Hopkins University Applied Physics Laboratory, Laurel, MD 20723, USA 6George Mason University, Fairfax, VA 22030, USA 7The Johns Hopkins University, Baltimore, MD 21218, USA 8Search for Extraterrestrial Intelligence Institute, Mountain View, CA 94043, USA 9Stanford University, Stanford, CA 94305, USA 10Nebraska Wesleyan University, Lincoln, NE 68504 11California Institute of Technology, Pasadena, CA 91125, USA 12Max-Planck-Institut für Sonnensystemforschung, 37191 Katlenburg-Lindau, Germany 13Groupe de Spectroscopie Moléculaire et Atmosphérique, Université Reims Champagne-Ardenne, 51687 Reims, France *To whom correspondence should be addressed.