Ecological and Economic Services Provided by Birds On
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Journal of Caribbean Ornithology
THE J OURNAL OF CARIBBEAN ORNITHOLOGY SOCIETY FOR THE C ONSERVATION AND S TUDY OF C ARIBBEAN B IRDS S OCIEDAD PARA LA C ONSERVACIÓN Y E STUDIO DE LAS A VES C ARIBEÑAS ASSOCIATION POUR LA C ONSERVATION ET L’ E TUDE DES O ISEAUX DE LA C ARAÏBE 2005 Vol. 18, No. 1 (ISSN 1527-7151) Formerly EL P ITIRRE CONTENTS RECUPERACIÓN DE A VES M IGRATORIAS N EÁRTICAS DEL O RDEN A NSERIFORMES EN C UBA . Pedro Blanco y Bárbara Sánchez ………………....................................................................................................................................................... 1 INVENTARIO DE LA A VIFAUNA DE T OPES DE C OLLANTES , S ANCTI S PÍRITUS , C UBA . Bárbara Sánchez ……..................... 7 NUEVO R EGISTRO Y C OMENTARIOS A DICIONALES S OBRE LA A VOCETA ( RECURVIROSTRA AMERICANA ) EN C UBA . Omar Labrada, Pedro Blanco, Elizabet S. Delgado, y Jarreton P. Rivero............................................................................... 13 AVES DE C AYO C ARENAS , C IÉNAGA DE B IRAMA , C UBA . Omar Labrada y Gabriel Cisneros ……………........................ 16 FORAGING B EHAVIOR OF T WO T YRANT F LYCATCHERS IN T RINIDAD : THE G REAT K ISKADEE ( PITANGUS SULPHURATUS ) AND T ROPICAL K INGBIRD ( TYRANNUS MELANCHOLICUS ). Nadira Mathura, Shawn O´Garro, Diane Thompson, Floyd E. Hayes, and Urmila S. Nandy........................................................................................................................................ 18 APPARENT N ESTING OF S OUTHERN L APWING ON A RUBA . Steven G. Mlodinow................................................................ -

Distribution, Ecology, and Life History of the Pearly-Eyed Thrasher (Margarops Fuscatus)
Adaptations of An Avian Supertramp: Distribution, Ecology, and Life History of the Pearly-Eyed Thrasher (Margarops fuscatus) Chapter 6: Survival and Dispersal The pearly-eyed thrasher has a wide geographical distribution, obtains regional and local abundance, and undergoes morphological plasticity on islands, especially at different elevations. It readily adapts to diverse habitats in noncompetitive situations. Its status as an avian supertramp becomes even more evident when one considers its proficiency in dispersing to and colonizing small, often sparsely The pearly-eye is a inhabited islands and disturbed habitats. long-lived species, Although rare in nature, an additional attribute of a supertramp would be a even for a tropical protracted lifetime once colonists become established. The pearly-eye possesses passerine. such an attribute. It is a long-lived species, even for a tropical passerine. This chapter treats adult thrasher survival, longevity, short- and long-range natal dispersal of the young, including the intrinsic and extrinsic characteristics of natal dispersers, and a comparison of the field techniques used in monitoring the spatiotemporal aspects of dispersal, e.g., observations, biotelemetry, and banding. Rounding out the chapter are some of the inherent and ecological factors influencing immature thrashers’ survival and dispersal, e.g., preferred habitat, diet, season, ectoparasites, and the effects of two major hurricanes, which resulted in food shortages following both disturbances. Annual Survival Rates (Rain-Forest Population) In the early 1990s, the tenet that tropical birds survive much longer than their north temperate counterparts, many of which are migratory, came into question (Karr et al. 1990). Whether or not the dogma can survive, however, awaits further empirical evidence from additional studies. -

The Effects of Hurricanes on Birds, with Special Reference to Caribbean Islands
Bird Conservation International (1993) 3:319-349 The effects of hurricanes on birds, with special reference to Caribbean islands JAMES W. WILEY and JOSEPH M. WUNDERLE, JR. Summary Cyclonic storms, variously called typhoons, cyclones, or hurricanes (henceforth, hurricanes), are common in many parts of the world, where their frequent occurrence can have both direct and indirect effects on bird populations. Direct effects of hurricanes include mortality from exposure to hurricane winds, rains, and storm surges, and geo- graphic displacement of individuals by storm winds. Indirect effects become apparent in the storm's aftermath and include loss of food supplies or foraging substrates; loss of nests and nest or roost sites; increased vulnerability to predation; microclimate changes; and increased conflict with humans. The short-term response of bird populations to hurricane damage, before changes in plant succession, includes shifts in diet, foraging sites or habitats, and reproductive changes. Bird populations may show long-term responses to changes in plant succession as second-growth vegetation increases in storm- damaged old-growth forests. The greatest stress of a hurricane to most upland terrestrial bird populations occurs after its passage rather than during its impact. The most important effect of a hurricane is the destruction of vegetation, which secondarily affects wildlife in the storm's after- math. The most vulnerable terrestrial wildlife populations have a diet of nectar, fruit, or seeds; nest, roost, or forage on large old trees; require a closed forest canopy; have special microclimate requirements and/or live in a habitat in which vegetation has a slow recovery rate. Small populations with these traits are at greatest risk to hurricane-induced extinction, particularly if they exist in small isolated habitat fragments. -

Bird Watcher's General Store
BIRD OBSERVER Carolina Wren © Barry Van Dusen, 1989 VOL. 18 NO. 6 DECEMBER 1990 BIRD OBSERVER VOL. 18 NO. 6 DECEMBER 1990 Editorial and Production Staff Dorothy R. Arvidson Theodore H. Atkinson Chere Bemelmans Associate Editor Brian E. Cassie Janet L. Heywood William E. Davis, Jr. Advisory Board Glenn d'Entremont Kathleen S. Anderson Herman H. D'Entremont James Baird H. Christian Floyd Alden G. Clayton Richard A. Forster Thomas W. French George W. Gove John C. Kricher Harriet E. Hoffman Ian C. T. Nisbet David E. Lange Bruce A. Sortie Simon A. Perkins Richeird K. Walton Wayne R. Petersen Martha Steele Corporate Officers Robert H. Stymeist William E. Davis, Jr., President Claudia Taylor Lee E. Taylor, Treasurer Lee E. Taylor H. Christian Floyd, Clerk Martha W. Vaughan BIRD OBSERVER{USPS 369-850) is published bimonthly, COPYRIGHT © 1990 by Bird Observer of Eastern Massachusetts, Inc., 462 Trapelo Road, Belmont, MA 02178, a nonprofit, tax-exempt corporation under section 501 (c)(3) of the Internal Revenue Code. Gifts to Bird Observer will be greatly appreciated and are tax deductible. POSTMASTER: Send address changes to BIRD OBSERVER, 462 Trapelo Road, Belmont, MA 02178. SUBSCRIPTIONS: $16 for 6 issues per calendar year, $30 for two years in the U. S. Add $2.50 per year for Canada and foreign. Single copies $3.00. An Index to Volumes 1-11 is $3. Back issues: inquire as to price and availabilify. CHANGES OF ADDRESS and subscription inquiries should be sent to Bird Observer Subscriptions, P. O. Box 236, Arlington, MA 02174. ADVERTISING: full page, $80; half page, $40; quarter page, $25. -

Avian Survey Report
Spring/Summer 2010 Avian Survey Report Stony Creek Wind Farm Wyoming County, New York January 24, 2011 PREPARED FOR: Stony Creek Energy LLC 51 Monroe St. Suite 1604 Rockville, MD 20850 PREPARED BY: Lackawanna Executive Park 239 Main Street, Suite 301 Dickson City, PA 18519 www.shoenerenvironmental.com Stony Creek Wind Farm Avian Survey January 24, 2011 Table of Contents I. Summary and Background .................................................................................................1 Summary .......................................................................................................................1 Project Description ........................................................................................................1 Project Review Background ..........................................................................................2 II. Bald Eagle Survey .............................................................................................................3 Bald Eagle Breeding Status in New York ......................................................................3 Daily Movements of Bald Eagle in New York ...............................................................4 Bald Eagle Conservation Status in New York ................................................................4 Bald Eagle Survey Method ............................................................................................5 Analysis of Bald Eagle Survey Data ..............................................................................6 -

Is Considered Globally Threatened Following the IUCN Red List Criteria
MEETING R EPORTS J. Carib. Ornithol. 18:88-93, 2005 TOWARDS A GLOBALLY THREATENED BIRD PROGRAM FOR THE CARIBBEAN 1 2 DAVID W EGE AND V ERÓNICA A NADÓN -IRIZARRY 1BirdLife International, Wellbrook Court, Girton Road, Cambridge CB3 0NA, UK; e-mail: [email protected]; 2Caribbean Program BirdLife International, Rio Canas 2107, Colorado St., Ponce, Puerto Rico 00728-1824; e-mail: [email protected] OVER 10% OF THE C ARIBBEAN ’S AVIFAUNA (57 3. Are there any priority actions that need to be of 560+ species) is considered globally threatened added to the published conservation action list for following the IUCN Red List criteria. Twelve of each species, and what level of funding would be these are Critically Endangered, 21 Endangered and required to implement the identified activities? 24 Vulnerable, with an additional 18 species Near 4. Would it be valuable for SCSCB to form a Threatened (see Table 1). These birds will be the Threatened Bird Working Group? next to go extinct (due to a number of differ- 5. What would be the focus of such a group, and ent threat factors), and thus it is our collective re- how would this relate to existing Working Groups? sponsibility to focus our attentions on their conser- vation. While conservation actions and monitoring Outputs .—The following outputs were discussed: activities are focused on some of these birds, for 1. There are 57 globally threatened and 18 near many we have no idea on a year to year basis threatened species on the IUCN Red List for the whether their status is improving or declining, or Caribbean (Table 1). -
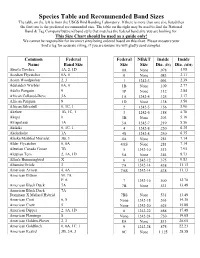
Species Table and Recommended Band Sizes the Table on the Left Is from the USGS Bird Banding Laboratory
Species Table and Recommended Band Sizes The table on the left is from the USGS Bird Banding Laboratory. If there is more than one size listed then the first one is the preferred recommended size. The table on the right may be used to find the National Band & Tag Company butt-end band style that matches the federal band size you are looking for. This Size Chart should be used as a guide only! We cannot be responsible for incorrect sizes being ordered based on this chart. Please measure your bird’s leg for accurate sizing, if you are unsure we will gladly send samples. Common Federal Federal NB&T Inside Inside Name Band Size Size Size Dia. (IN) Dia. (MM) Abert's Towhee 1A, 2, 1D 0A None .078 1.98 Acadian Flycatcher 0A, 0 0 None .083 2.11 Acorn Woodpecker 2, 3 1 1242-3 .094 2.39 Adelaide's Warbler 0A, 0 1B None .109 2.77 Adelie Penguin 9 1P None .112 2.84 African Collared-Dove 3A 1A 1242-4 .125 3.17 African Penguin 9 1D None .138 3.50 African Silverbill 0, 1C, 1 2 1242-5 .156 3.96 Akekee 1B, 1C, 1 3 1242-6 .188 4.78 Akepa 0 3B None .203 5.16 Akiapolaau 1A 3A 1242-7 .219 5.56 Akikiki 0, 1C, 1 4 1242-8 .250 6.35 Akohekohe 1A 4S 1242-8 .250 6.35 Alaska Marbled Murrelet 3B, 3 4A None .281 7.14 Alder Flycatcher 0, 0A 4AS None .281 7.14 Aleutian Canada Goose 7B 5 1242-10 .313 7.95 Aleutian Tern 2, 1A, 1D 5A None .344 8.73 Allen's Hummingbird X 6 1242-12 .375 9.53 Altamira Oriole 3 7A 1242-14 .438 11.13 American Avocet 4, 4A 7AS 1242-14 .438 11.13 American Bittern M: 7A F: 6 7 1242-16 .500 12.70 American Black Duck 7A 7B None .531 13.49 American -

Ecological and Economic Services Provided by Birds on Jamaican Blue Mountain Coffee Farms
Contributed Paper Ecological and Economic Services Provided by Birds on Jamaican Blue Mountain Coffee Farms JHERIME L. KELLERMANN,∗‡ MATTHEW D. JOHNSON,∗ AMY M. STERCHO,∗ AND STEVEN C. HACKETT† ∗Department of Wildlife, Humboldt State University, Arcata, CA 95521, U.S.A. †Department of Economics, Humboldt State University, Arcata, CA 95521, U.S.A. Abstract: Coffee farms can support significant biodiversity, yet intensification of farming practices is de- grading agricultural habitats and compromising ecosystem services such as biological pest control. The coffee berry borer (Hypothenemus hampei) is the world’s primary coffee pest. Researchers have demonstrated that birds reduce insect abundance on coffee farms but have not documented avian control of the berry borer or quantified avian benefits to crop yield or farm income. We conducted a bird-exclosure experiment on coffee farms in the Blue Mountains, Jamaica, to measure avian pest control of berry borers, identify potential predator species, associate predator abundance and borer reductions with vegetation complexity, and quan- tify resulting increases in coffee yield. Coffee plants excluded from foraging birds had significantly higher borer infestation, more borer broods, and greater berry damage than control plants. We identified 17 poten- tial predator species (73% were wintering Neotropical migrants), and 3 primary species composed 67% of migrant detections. Average relative bird abundance and diversity and relative resident predator abundance increased with greater shade-tree cover. Although migrant predators overall did not respond to vegetation complexity variables, the 3 primary species increased with proximity to noncoffee habitat patches. Lower infes- tation on control plants was correlated with higher total bird abundance, but not with predator abundance or vegetation complexity. -

Fernando Marques Dos Santos BIOLOGIA REPRODUTIVA DE PASSERIFORMES SUBTROPICAIS DO SUL DO BRASIL
Fernando Marques dos Santos BIOLOGIA REPRODUTIVA DE PASSERIFORMES SUBTROPICAIS DO SUL DO BRASIL: testando a teoria de convergência latitudinal das fenologias reprodutivas Dissertação apresentada ao Setor de Ciências Biológicas da Universidade Federal do Paraná, como parte dos requisitos para a obtenção do grau de mestre em Ecologia e Conservação. Orientador: Dr. James Joseph Roper Curitiba – PR 2014 AGRADECIMENTOS À Uschi Wischhoff pelos anos de harmonia entre colaboração científica e convivência. Ao James Roper pelos valiosos conselhos e incentivo em observar a natureza. À Talita Braga, Gustavo Cerboncini e Rafaela Bobato pelo auxílio em campo de extrema competência. À Bióloga Ana Cristina Barros pelo bom humor e receptividade nas áreas de estudo. Aos guardas dos Mananciais da Serra e Casa da Cultura da Água pelo respeito à fauna, e especialmente Odirlei, Emerson e Carlos pelos ninhos encontrados. Aos professores e funcionários da PPGECO pela imensurável dedicação. À CAPES pela bolsa de mestrado. Aos meus pais, Ocimara e Mário, por tudo. Ao povo brasileiro pela oportunidade em contribuir com conhecimento de importância à longo prazo. “Every now and then I break free from the office, the computer, the telephone, the piles of manuscripts, and take up an invitation to visit a field site somewhere. I stumble over logs, get ripped by thorns, bitten by horseflies, stuck in mud, sunburned, and bruised. I sweat, groan, spit, curse, and generally have a wonderful time. I fight back tears when I see old, long- forgotten friends---the wildflowers, ferns, trees, salamanders, fungi, and beetles I once knew so well but whose names now elude me as often as not. -

HISPANIOLA & PUERTO RICO with JAMAICA & the BAHAMAS
Broad-billed Tody (Eustace Barnes) HISPANIOLA & PUERTO RICO with JAMAICA & THE BAHAMAS 29 MARCH/5 APRIL – 17/22 APRIL 2015 LEADER: EUSTACE BARNES 1 BirdQuest Tour Report: Hispaniola & Puerto Rico with Jamaica & the Bahamas 2015 www.birdquest-tours.com The Gang (most of) at Cabo Rojo on Puerto Rico (Eustace Barnes) A glorious mix of blue skies, verdant rainforests, thorny woodlands, lush marshes, red sandstone cliffs, mangroves and azure seas greeted us on each and every day as we toured through the Greater Antilles and Bahamas seeing all but one endemic on our way. Jamaica is truly a gem and staying at Marshall’s Pen a real pleasure with Jamaican Owls in the grounds and Red-billed Streamertails buzzing about. High above Kingston, Crested Quail Doves before breakfast followed by Blue Mountain Coffee takes some beating as does watching Black-billed Streamertails at a lek. The Dominican Republic was more demanding but gave us some great moments including La Selle Thrush, Hispaniolan Trogon and a singing White-fronted Quail Dove within the space of a few minutes of one another. Our forest camp gave us another White-fronted Quail Dove and Key West Quail Dove running about in front of us just before we found a Least Poorwill! An Ashy- faced Owl gave unbeatable views and our search for Ridgway’s Hawk was dramatically shortened when a female landed in an open tree in front of us. Puerto Rico is a truly American Experience providing car park birding at its best. The Elfin Woods Warbler and Puerto Rican Screech Owl car park experience was the best. -

Jamaica Island Endemics II 12Th – 17Th March 2017 (6 Days) Trip Report
Jamaica Island Endemics II 12th – 17th March 2017 (6 Days) Trip Report Red-billed Streamertail by David Howe Trip Report compiled by tour leader, Forrest Rowland Trip Report – RBL Jamaica - Island Endemics II 2017 2 ___________________________________________________________________________________ Tour Top 5: 1. Crested Quail Dove 4. Jamaican Lizard Cuckoo 2. Red-billed Streamertail 5. Northern Potoo 3. Jamaican Tody Tour Summary Jamaica is as about unique a destination as can be imagined. In the middle of the Caribbean, this small island developed a culture unlike any other, cuisine unlike any other, and vibrant, beautiful avifauna unlike any other. As a guide, it’s pretty dreamy as well. Great accommodations, great food, and outstanding local bird guides that are extremely knowledgeable and helpful, making it a delight for everyone involved! Our group met up in Montego Bay this year, rather than Kingston. With more international flights available to this extremely touristy destination, schedules were more suited to this port, though it did mean missing a chance in the Hellshire Hills for the Bahama Mockingbird. Luckily, we made it up for it with other amazing species and encounters later in the tour. After a brief lunch stop en route, we made our way straight through to the lovely Greencastle Estate for some evening birding, picking up Glossy Ibis, Neotropic Cormorant, Laughing Gull, and a few Jamaican Euphonia by David Howe other commoner species before we had arrived. Greencastle Estate is situated just inland from the seashore. Perched atop a hill with an incredible view of the Blue Mountains, Annotto Bay, and the Caribbean, this old plantation home was renovated and converted into a finely-appointed Lodge, complete with swimming pool, tennis court, and gardens. -

Jamaica January, 2018
Tropical Birding Trip Report JAMAICA JANUARY, 2018 A Tropical Birding CUSTOM TOUR Jamaica: The Caribbean Introtour 11-16 January, 2018 TOUR LEADER: Charley Hesse Local guide: Ricardo Miller Report & photos by Charley Hesse. All photos were taken on this tour The undeniable star of the show: The Jamaican Tody is a little gem. Jamaica is the perfect place to begin exploring the Caribbean. It is easily reached from the US with regular flights, there is a good tourist infrastructure, and the impressive crop of endemics (27 by a conservative count) tops the Carribean. The birding is relatively easy, and one has a good chance of seeing every endemic and even photographing many of them. The charm of the friendly locals, a distinctive cuisine, fantastic music, stunning scenery and world-famous coffee all add extra reasons to visit this already attractive destination. It’s not just number of endemics either. The list boasts some absolute corkers with birds like Jamaican Tody, the long-tailed Streamertail hummingbird and Jamaican Owl being perennial favorites. Lush montane forest in the famous Blue Mountains National Park holds the vast majority of Jamaica’s endemics, but dry forest of the Hellshire Hills, relaxing botanical gardens, the famous Ecclesdown Road in the base of the John Crow Mountains, and even a splash of birding on the coast and wetlands gives a very varied trip. (Although this was a custom trip, it follows the same itinerary as our set departure tour.) www.tropicalbirding.com +1-409-515-9110 [email protected] Tropical Birding Trip Report JAMAICA JANUARY, 2018 12th January – Kingston to the Blue Mountains After a late arrival last night, we started our birding in the hotel parking lot this morning, while we waited for our local guide to arrive.