The Pleistocene of Simila Open Pit (Scythian Platform, Romania)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Nr. Crt Tipul Activitatii Subiectul Abordat Data/Perioad a Desfasurarii
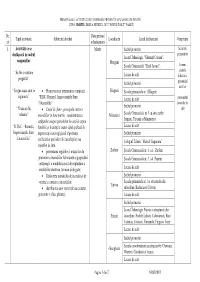
PRINCIPALELE ACTIVITATI DE INFORMARE PREVENTIVA PLANIFICATE PENTRU LUNA MARTIE 2013 LA NIVELUL ISUJ “PODUL ÎNALT” VASLUI Nr. Data/perioad Tipul activitatii Subiectul abordat Localitatea Locul desfasurarii Grup tinta crt a desfasurarii 1. Activităţi ce se Martie Sediul primariei Salariatii desfăşoară in cadrul primariilor Liceul Tehnologic “Ghenuţă Coman”, campaniilor Murgeni Şcoala Gimnazială “Emil Juvara”, liceeni, “Să fim o naţiune cadrele Lacase de cult didactice, pregătită” Sediul primariei personalul auxiliar “ Singur acasa sunt in • Promovarea şi prezentarea campaniei Blagesti Scoala gimnaziala nr 1 Blagesti siguranta” “RISC- Renunţă, Improvizaţiile Sunt Lacase de cult personalul Catastrofale” lacaselor de Sediul primariei “ Vreau sa fiu • Coşul de fum – principala cauza a cult voluntar” incendiilor in luna martie conştientizarea Malusteni Şcoala Gimnazială nr 1 cu structurile: cetăţenilor asupra pericolelor la care îşi expun Lupesti, Tutcani si Manzatesti “R.I.S.C. –Renunţă, familiile şi locuinţele atunci când apelează la Lacase de cult Improvizaţiile Sunt improvizaţii sau neglijează importanţa Sediul primariei Catastrofale” verificărilor periodice ale instalaţiilor sau Colegiul Tehnic “Marcel Guguianu” coşurilor de fum. • prezentarea regulilor şi măsurilor de Zorleni Şcoala Gimnazială nr. 1, sat. Zorleni prevenire a incendiilor la locuinţe şi gospodării Şcoala Gimnazială nr. 3, sat. Popeni cetăţeneşti, a modului corect de exploatare a instalaţiilor electrice, termice şi de gaze; Lacase de cult • Reducerea numărului de incendii -

O R D I N U L Nr. 498 Privind Numerotarea Secţiilor De Votare Din
O R D I N U L Nr. 498 privind numerotarea secţiilor de votare din judeţul Vaslui pentru alegerea membrilor din România în Parlamentul European din 25 noiembrie 2007 Ciprian Iftimoaei - Prefectul judeţului Vaslui ; având în vedere dispoziţiile art. 25 alin. 1 din Legea nr. 373/2004 privind alegerea Camerei Deputaţilor şi a Senatului, republicată, art. 16, art.17 alin. 1 şi 2 din Legea nr. 33/2007 privind organizarea şi desfăşurarea alegerilor pentru Parlamentul European, cu modificările şi completările ulterioare şi ale Ordonanţei de Urgenţă nr. 15/2007 privind unele măsuri referitoare la alegerea membrilor României în Parlamentul European în anul 2007; în temeiul art. 32 din Legea nr. 340/2004 privind instituţia prefectului, cu modificările şi completările ulterioare, emit prezentul O R D I N : Art. 1. - Se delimitează şi numerotează secţiile de votare din judeţul Vaslui pentru alegerea membrilor din România în Parlamentul European din 25 noiembrie 2007, conform anexei care face parte integrantă din prezentul Ordin. Art. 2. Delimitarea şi numerotarea fiecărei secţii de votare de pe raza judeţului Vaslui se aduce la cunoştinţă cetăţenilor prin PUBLICAŢIE în care se vor indica şi locurile de desfăşurare a votării. Dat astăzi, 24 septembrie 2007 P R E F E C T , SUBPREFECT, Ciprian Iftimoaei Vasile Cupşan - Cătălin AVIZAT FAVORABIL, Direcţia Verificarea Legalităţii Actelor a Aplicării Actelor Normative şi Contencios Administrativ, D I R E C T O R, Costică Coatu Întocmit, Costică Coatu 25.09.2007-3 ex. AA DELIMITAREA ŞI NUMEROTAREA SECŢIILOR DE VOTARE DIN JUDEŢUL VASLUI Secţia de votare nr. 1 – din str. Ştefan cel Mare nr. -

Vaslui County in the National Context: Evolution and Effects of Mobility of the Working Age Population
A Service of Leibniz-Informationszentrum econstor Wirtschaft Leibniz Information Centre Make Your Publications Visible. zbw for Economics Munteanu, Roxana Florina Article Vaslui county in the national context: Evolution and effects of mobility of the working age population CES Working Papers Provided in Cooperation with: Centre for European Studies, Alexandru Ioan Cuza University Suggested Citation: Munteanu, Roxana Florina (2017) : Vaslui county in the national context: Evolution and effects of mobility of the working age population, CES Working Papers, ISSN 2067-7693, Alexandru Ioan Cuza University of Iasi, Centre for European Studies, Iasi, Vol. 9, Iss. 2, pp. 123-138 This Version is available at: http://hdl.handle.net/10419/198502 Standard-Nutzungsbedingungen: Terms of use: Die Dokumente auf EconStor dürfen zu eigenen wissenschaftlichen Documents in EconStor may be saved and copied for your Zwecken und zum Privatgebrauch gespeichert und kopiert werden. personal and scholarly purposes. Sie dürfen die Dokumente nicht für öffentliche oder kommerzielle You are not to copy documents for public or commercial Zwecke vervielfältigen, öffentlich ausstellen, öffentlich zugänglich purposes, to exhibit the documents publicly, to make them machen, vertreiben oder anderweitig nutzen. publicly available on the internet, or to distribute or otherwise use the documents in public. Sofern die Verfasser die Dokumente unter Open-Content-Lizenzen (insbesondere CC-Lizenzen) zur Verfügung gestellt haben sollten, If the documents have been made available -

Nr.820 /09.05.2012 SITUAŢIA CENTRALIZATOARE A
MINISTERUL AGRICULTURII ŞI DEZVOLTĂRII RURALE DIRECŢIA PENTRU AGRICULTURĂ A JUDETULUI VASLUI Serviciul Inspecţii,Implementare Politici de Piaţă Str. Eternităţii, nr. 1, Vaslui Telefon: 0040235311802, Fax: 0040235311591 E-mail: [email protected]; wwwdirectiagricolavs.ro Nr.820 /09.05.2012 SITUAŢIA CENTRALIZATOARE A OPERATORILOR ECONOMICI DIN JUDEŢUL VASLUI CARE EXPLOATEAZĂ SPAŢII DE DEPOZITARE Date identificare spațiu Capacitatea de depozitare aflat în Din care Nr. Autorizația de Date identificare operator totală Tipul instalatiei depozit exploatare Crt. economic autorizată de cantarire (serie, număr) (denumire, adresă, telefon, e-mail) (denumire, adresă, Siloz Magazie (tone) telefon) (tone) (tone) VS.0000601/2006 SC. MÂNDRA.SA ‐BÂRLAD 4000 2000 2000 Unit. depozit. Str. Mihai Viteazul nr.37,loc. dispozitiv de SC. MÂNDRA.SA BÂRLAD Bârlad, cântărire care Str. Mihai Viteazul nr.37 jud. Vaslui,Tel.0235/421070, eliberează în mod Loc. Bârlad, jud. Vaslui E‐mail automat tichetul de 1 Tel.0235/421070, ;[email protected] cântar VS.0000602/2006 8000 3000 5000 SC.VECAST S.R.L. BÂRLAD dispozitiv de SC.VECAST S.R.L. BÂRLAD Unit. depozit. cântărire care Str. Palerma nr. 4 Str.Palerma nr. 4,loc. Bârlad, eliberează în mod Loc. Bârlad, jud. Vaslui jud. Vaslui automat tichetul de 2 Tel .0722326595 Tel .0722326595 cântar VS.0000603/2006 SC.AGROMEC ZORLENI SA 1250 0 1250 Unit.depozit. dispozitive de cântărire SC.AGROMEC ZORLENI SA Loc. Zorleni, com. Zorleni, pentru care tichetul de Loc. Zorleni,Com. Zorleni, Jud. Vaslui Jud. Vaslui cântar se completeaza 3 Tel. 0235/424898 Tel. 0235/424898 de operatorul economic VS.0003481/2009 SC.ULEROM. SA VASLUI 20600 3000 Unit. depozit. -

CABINETE MEDICINA DE FAMILIE LA 23.04.2019 1CM! Med. Fam. Dr
CABINETE MEDICINA DE FAMILIE LA 23.04.2019 1CM! Med. Fam. Dr. ABABEI LILIANA Vaslui, str. Hagi Chiriae, nr. 56, 0235312703 2 CM! Med. Fam. Dr. APOSTOL MIHAELA Vaslui, str. Dimitrie Bolintineanu, or. 10, 0235314882 3 CM! Med. Fam. Dr. ARDELEANU ECATERINA Vaslui, str. Dimitrie Bolintineanu, or. 10 0235314824 4 CMI Med. Fam. Dr. BARLADEANU RALUCA Vaslui, str. Republieii, bL351 0235360101 5 CM! Med. Fam. Dr. BAZDARA MIMI Vaslui, str. Bueuresti, bI.410, se.a, ap. 1 0335404468 Vaslui, STR. TRAIAN, BL. 241, SC. E, 6 CMI Med. Fam. Dr. BUHUS LIGIA NICOLETA P, AP. 21 0733018551 7 CMI Med. Fam. Dr. CRISTEA CARMEN DOINA Vaslui, str.Avantului,48 0753034966as;0747671373 8 CMI Med. Fam. Dr. DAMIAN DANIELA TEODORA Vaslui, str. Republieii, bI.351 0335412045 9 CMI Med. Fam. Dr. DUMITRIU ALINA Vaslui, str. Republieii, bI.351 0235316950 CM! Med. Fam. Dr. GENTIMIR LIBELULA - eu pet. 10 Lueru Deleni Vaslui, Str. Doniei, bl. 21, ap. 19 0235315366 Vaslui, str.Vasile Aleesandri, bI.31, CMI Med. Fam. Dr. GHERAN CONSTANTIN 11 ap.19, parter, 0235313133 12 CMI Med. Fam. Dr. GRECU IONEL CRISTINEL Vaslui, Str. Paeii, bi. 362, se. A, ap. 17 0235311638 13 CMI Med. Fam. Dr. GUGIUMAN MARILENA Vaslui, str. Dimitrie Bolintineanu,or. 10 0235311572 CMI Med. Fam. Dr. LACATUSU LAURA MONICA 14 RODICA Vaslui, str. Republieii, bI.351 0235360223 15 CMI Med. Fam. Dr. NIMIRCEAG MARIUSS Vaslui, str. Spiru Haret, or.1 0235369368 16 CM! Med. Fam. Dr. OUATV GABRIELA Vaslui, str. Dimitrie Bolintineanu, or. 10 0741180183 17 CMI Med. Fam. IGNATESCU LIDIA Vaslui, str. Dimitrie Bolintineanu,or. -

Tabelul Cu Persoanele Care Au Donat Pentru Construirea Şi Amplasarea Troiţei În Incinta Casei De Ajutor Reciproc ,,Elena Cuza
Tabelul cu persoanele care au donat pentru construirea şi amplasarea troiţei în incinta Casei de Ajutor Reciproc ,,Elena Cuza" Bârlad Nr Nume şi prenume Suma Donaţie crt 1 S.C. An&An Consult - Grăjdeni prin Ciobanu Andrei 2.000 Troiţă 2 S.C. AMC VECTOR S.R.L. Bârlad prin Hriscu Vlad Andrei 1.000 Troiţă 3 S.C. TRITON S.R.L. Iaşi prin Cocea Viorel 1.000 Troiţă 4 S.C. CRISMIH Bârlad prin Mihăilă Cristinel 1.000 Troiţă 5 S.C. NEXTEL INVEST S.R.L. Focşani prin Pancu Daniel 1.000 Troiţă 6 CMI Dr ICHIM LAURENŢIA ADRIANA - Bârlad 500 Troiţă 7 S.C. CORMORAN S.R.L. Vaslui prin Solomon Nicolae 400 Troiţă 8 S.C. TIPARUL S.R.L. Bârlad prin Stoian Neculai 300 Troiţă 9 S.C. 3D BIOSERVSAN S.R.L. Bârlad prin Catanoiu Mugurel 220 Troiţă 10 S.C. GAMISOFT S.R.L. Bârlad prin Silion Gabi 200 Troiţă 11 S.C. NEGOCOMIS S.R.L Bârlad prin Gîfu Gheorghe 200 Troiţă 12 CMI Dr IVAN AURA ANDREEA - Bârlad 200 Troiţă 13 P.F.A. Doctor COJOCARU LUCIAN - Bârlad 100 Troiţă 14 S.C. VERTRANTIS S.R.L. prin Rusu Mihaela - Bârlad 100 Troiţă 15 S.C. ELECTOINST SCÎNTEIE - Bârlad prin Scînteie Cezar Gabriel 52 Troiţă 16 CMI Dr DORNIANU RODICA - Bârlad 50 Troiţă 17 Lovin Alexandra - Sat Popeni, Comuna Zorleni 1.350 Troiţă 18 Dima Dumitru - Bârlad 1.100 Troiţă 19 Mihai Nicolaie - Bârlad 1.100 Troiţă 20 Cozma Cristian Cătălin - Bârlad 1.050 Troiţă 21 Clime Ionel - Bârlad 1.000 Troiţă 22 Mihai Lenuţa - Bârlad 900 Troiţă 23 Munteanu Luminiţa - Bârlad 850 Troiţă 24 Rîpan Rodica - Bârlad 780 Troiţă 25 Hriscu Vlad Andrei - Bârlad 700 Troiţă 26 Râncu Liviu Adrian - Bârlad 700 Troiţă 27 Rîpan -
The Pleistocene of Simila Open Pit (Scythian Platform, Romania)
Muzeul Olteniei Craiova. Oltenia. Studii şi comunicări. Ştiinţele Naturii. Tom. 29, No. 1/2013 ISSN 1454-6914 THE PLEISTOCENE OF SIMILA OPEN PIT (SCYTHIAN PLATFORM, ROMANIA) CODREAVlad, RĂȚOI G. Bogdan, URSACHI Laurențiu, SOLOMON Alexandru, BRÂNZILĂ Mihai Abstract. Pleistocene deposits recorded on the Bârlad Valley area are largely exposed, mainly on the river terraces. An illustrative outcrop can be observed on the right bank of the Bârlad River few kilometres upstream from Bârlad town, in Simila open pit. Based on detailed study, a number of characteristic sedimentary packages were outlined, referred to as lithofacies associations. There were established a total of eight lithofacies types and three architectural elements. The interpretation of these architectural elements revealed a braided river environment with low sinuosity channels. There are two kinds of fossils collected in the sands and gravels from Simila: Late Miocene ones (fossil wood, molluscs, and vertebrates), all being reworked there; Pleistocene (most probably Middle or Late Pleistocene) ones, including freshwater molluscs and vertebrates (large herbivores). Therefore, the deposits from Simila open pit are Pleistocene and in no wise, older as presumed before. Keywords: Pleistocene, fluvial environments, sedimentology, palaeontology, Bârlad Valley. Rezumat. Pleistocenul din cariera Simila (Platforma Scitică, România). În aria Văii Bârladului, depozite pleistocene sunt consemnate îndeosebi la nivelurile diferitelor terase. Un afloriment deosebit de ilustrativ poate fi studiat pe malul drept al râului Bârlad, la câțiva kilometri amonte de orașul Bârlad, în cariera de la Simila. Pe baza studiului detaliat efectuat, au fost conturate o serie de litofaciesuri sedimentare grupate unor elemente arhitecturale. Au fost astfel stabilite opt tipuri de litofaciesuri sedimentare și trei elemente arhitecturale. -

Activităţile Metodico-Ştiinţifice Şi Psihopedagogice
MINISTERUL EDUCAŢIEI NAŢIONALE INSPECTORATUL ŞCOLAR JUDEŢEAN VASLUI * CASA CORPULUI DIDACTIC * Strada Ştefan cel Mare, nr. 37, Vaslui, Cod 730178 Telefon: 0235 317279, 0335402273 Fax : 0235/31.72.79 e-mail: [email protected], [email protected] http://www.ccdvaslui.ro Avizat, Inspector Şcolar General, Prof. GABRIELA PLĂCINTĂ Director, Casa Corpului Didactic Vaslui Prof. CRISTINEL POPA organizate pentru personalul didactic şi didactic auxiliar din învăţământul preuniversitar =2= SEPTEMBRIE OCTOMBRIE NOIEMBRIE DECEMBRIE L M M J V S D L M M J V S D L M M J V S D L M M J V S D 1 1 2 3 4 5 6 1 2 3 1 2 3 4 5 6 7 8 7 8 9 10 11 12 13 4 5 6 7 8 9 10 2 3 4 5 6 7 8 9 10 11 12 13 14 15 14 15 16 17 18 19 20 11 12 13 14 15 16 17 9 10 11 12 13 14 15 21 22 23 24 25 26 27 16 17 18 19 20 21 22 18 19 20 21 22 23 24 16 17 18 19 20 21 22 23 24 25 26 27 28 29 28 29 30 31 23 24 25 26 27 28 29 25 26 27 28 29 30 30 30 31 S M T W T F S S M T W T F S S M T W T F S S M T W T F S IANUARIE FEBRUARIE MARTIE APRILIE L M M J V S D L M M J V S D L M M J V S D L M M J V S D 1 2 3 4 5 1 2 1 2 1 2 3 4 5 6 6 7 8 9 10 11 12 3 4 5 6 7 8 9 3 4 5 6 7 8 9 13 14 15 16 17 18 19 7 8 9 10 11 12 13 10 11 12 13 14 15 16 10 11 12 13 14 15 16 20 21 22 23 24 25 26 14 15 16 17 18 19 20 17 18 19 20 21 22 23 17 18 19 20 21 22 23 27 28 29 30 31 21 22 23 24 25 26 27 24 25 26 27 28 24 25 26 27 28 29 30 28 29 30 31 S M T W T F S S M T W T F S S M T W T F S S M T W T F S MAI IUNIE IULIE AUGUST L M M J V S D L M M J V S D L M M J V S D L M M J V S D 1 2 3 4 1 1 2 3 4 5 6 1 2 3 5 6 7 8 9 10 11 2 3 4 -

Plan De Şcolarizare Clasa Pregătitoare an Şcolar 2021-2022
Plan de şcolarizare Clasa pregătitoare An şcolar 2021-2022 Masă / Normal / Step by Step / Buget / Limba română Număr clase Nr. crt. Denumire unitate Localitate Localitate superioară Capacitate (locuri) aprobate 1 ȘCOALA GIMNAZIALĂ "EPISCOP IACOV ANTONOVICI", MUN. BÂRLAD BÂRLAD MUNICIPIUL BÂRLAD 1 30 2 ŞCOALA GIMNAZIALĂ "MIHAIL SADOVEANU", MUN. HUŞI HUŞI MUNICIPIUL HUŞI 1 25 3 ȘCOALA GIMNAZIALĂ "ALEXANDRA NECHITA", MUN. VASLUI VASLUI MUNICIPIUL VASLUI 1 25 4 ȘCOALA GIMNAZIALĂ "CONSTANTIN PARFENE", MUN. VASLUI VASLUI MUNICIPIUL VASLUI 1 25 Total Masă / Normal / Step by Step / Buget / Limba română 4 105 Pagina 1 / 12 Masă / Normal / Tradițional / Buget / Limba română Număr clase Nr. crt. Denumire unitate Localitate Localitate superioară Capacitate (locuri) aprobate 1 ȘCOALA GIMNAZIALĂ NR. 1, SAT GARA BANCA GARA BANCA BANCA 1 17 2 ȘCOALA GIMNAZIALĂ NR. 1, SAT BĂCANI BĂCANI BĂCANI 1 21 3 ȘCOALA GIMNAZIALĂ "TEODOR JUNCU", SAT BĂCEȘTI BĂCEŞTI BĂCEŞTI 2 40 4 ȘCOALA GIMNAZIALĂ "ANASTASIE FĂTU", SAT BEREZENI BEREZENI BEREZENI 2 30 5 ȘCOALA GIMNAZIALĂ "HORIA STAMATIN", SAT BOGDĂNEȘTI / ȘCOALA GIMNAZIALĂ, SAT UNȚEŞTI BOGDĂNEŞTI 1 10 UNȚEȘTI 6 ȘCOALA GIMNAZIALĂ NR. 1, SAT AVEREȘTI / ȘCOALA GIMNAZIALĂ, SAT BUNEȘTI BUNEŞTI BUNEŞTI-AVEREŞTI 1 12 7 LICEUL "ȘTEFAN CEL MARE", SAT CODĂEȘTI CODĂEŞTI CODĂEŞTI 1 15 8 ȘCOALA GIMNAZIALĂ "SFÂNTUL IERARH LUCA", SAT COSTEȘTI COSTEŞTI COSTEŞTI 1 14 9 ȘCOALA GIMNAZIALĂ "CEZAR BOTEZ", SAT FÂSTÂCI FÂSTÂCI COZMEŞTI 1 21 10 ȘCOALA GIMNAZIALĂ NR. 1, SAT DELEȘTI DELEŞTI DELEŞTI 1 10 11 ȘCOALA GIMNAZIALĂ "VICTOR ION POPA", SAT DODEȘTI / ȘCOALA PRIMARĂ TĂMĂȘENI, DODEŞTI DODEŞTI 1 14 SAT DODEȘTI 12 ȘCOALA GIMNAZIALĂ NR. 1, SAT RĂDENI RĂDENI DRAGOMIREŞTI 2 28 13 ȘCOALA GIMNAZIALĂ "PROF. -

Geological Study of Husi – Murgeni – Bârlad – Vaslui – Muntenii De Sus Area
”ALEXANDRU IOAN CUZA” UNIVERSITY IAŞI FACULTY OF GEOGRAPHY AND GEOLOGY DEPARTMENT OF GEOLOGY DOCTORAL SCHOOL GEOLOGICAL STUDY OF HUSI – MURGENI – BÂRLAD – VASLUI – MUNTENII DE SUS AREA SUMMARY -DOCTORATE THESIS- Scientific Coordinator: Prof. univ. dr. Mihai BRÂNZILĂ PhD student: BEJAN Daniel IAŞI 2012 "ALEXANDRU IOAN CUZA" UNIVERSITY IAŞI RECTORATE Mr./Mrs. ………………………………………. On the 29th of September 2012, at 10 o’clock, in the amphitheater B6, at the Faculty of Geography and Geology, the Geology Department, Mr. Bejan Daniel will give a presentation, in open session, of his doctorate thesis entitled ”GEOLOGICAL STUDY OF HUȘI – MURGENI – BÂRLAD – VASLUI – MUNTENII DE SUS AREA” for obtaining the scientific title of doctor in Geology. The doctorate committee is made up of: President: Associate Professor, PhD Traian GAVRILOAIEI – deputy dean of the Faculty of Geography and Geology,”Al. I. Cuza” University, Iaşi. Scientific coordinator: University Professor, PhD Mihai BRÂNZILĂ - the Faculty of Geography and Geology, Al. I. Cuza” University, Iaşi. Reviewers: University Professor, PhD Vlad A. CODREA, Faculty of Biology and Geology, The Geology Department, ”Babeş-Bolyai” University, Cluj-Napoca PhD Titus BRUSTUR, Scientific researcher 1st degree, The National Marine Geology and Geo-ecology Institute (GEOECOMAR), Bucharest. Associate Professor PhD Paul ŢIBULEAC, Faculty of Geography and Geology, The Geology Department, ”Al. I. Cuza” University, Iaşi. You are invited to attend the session for the presentation of the thesis. The thesis can be read -

Autoritatea Naţională Sanitară Veterinară Şi Pentru Siguranţa Alimentelor
AUTORITATEA NAŢIONALĂ SANITARĂ VETERINARĂ ŞI PENTRU SIGURANŢA ALIMENTELOR DIRECŢIA SANITARĂ VETERINARĂ ŞI PENTRU SIGURANŢA ALIMENTELOR VASLUI Nr. CIRCUMSCRIPTIA CONCESIONAR/PRESTATOR NUME SI PRENUME crt. SANITAR- CONCESIONAR/PRESTATOR DATE DE VETERINARA DE CONTACT(ADRESA, EMAIL, NR.DE ASISTENTA TELEFON) 1. AL. VLAHUTA SC FLORIVASVET SRL Dr. Olenici Ifrim, loc. Buda,com. Al.Vlahuta, tel. 0766271875, [email protected] 2. ARSURA SC HELASMED SRL Dr. Hapenciuc Elena, loc. Ghermanesti, com.Dranceni, tel.0766723865, [email protected] 3. AVERESTI SC SANTEANIMAVET SRL Dr. Balu Ciprian, loc. Satu Nou, com. Muntenii de Sus, tel.0744841151, [email protected] 4. BACANI SC PROVET PLETEA SRL Dr. Pletea Mihai Catalin, loc. Bacani, com.Bacani, tel.0763978640, 5. BĂCESTI CMVI ŢIGU ILIE Dr. Tigu Ilie, loc. Bacesti, com. Bacesti, tel.0740842414, [email protected] 6. BALTENI SC BIOROMAD SRL Dr.Rotaru Florin, Loc.Balteni, com.Balteni, tel.0740484792, [email protected] 7. BANCA SC VANBET SRL Dr.Bogos Fanel, loc. Salcioara nr. 15, com.Banca, tel.0765265577, [email protected] Pagina 1 din 8 Adresa: Barlad, str. Trestiana, nr.2, jud. Vaslui Cod Poştal 731030; Telefon: 0235421121, 0235421413, Fax: 0235421278 E-mail: [email protected] Web: www.ansvsa.ro. 8. BARLAD SC MEDI INTERVET SRL Dr.Bejan Ioan, Str, stefan cel Mare nr. 56, mun. Barlad, tel.0740256913, [email protected] 9. BEREZENI SC MEDVICOL SRL Dr.Vicol Gheorghe, loc. Berezeni ,nr.62, com. Berezeni, tel.0762652153, [email protected] 10. BLĂGEŞTI CMVA DR.CAIN ANTON Dr. Cain Anton, Str. Emil Juvara, nr. 618, oras Murgeni, tel.0744436370, [email protected] 11. BOGDANA SC FAMAGROMED SRL Dr. -

Județul Vaslui
39DILúDUHMXGHĠ Proces-verbal din data 24.05.2016SULYLQGGHVHPQDUHDSUHúHGLQĠLORUELURXULORUHOHFWRUDOHDOHVHFĠLLORUGHYRWDUH constituite pentru alegerile locale din anul 2016úLDORFĠLLWRULORUDFHVWRUD &LUFXPVFULSĠLD(OHFWRUDOă-XGHĠHDQăQU39, VASLUI ,QL܊LDOD Nr. crt. UAT 1U6HF܊LH ,QVWLWX܊LD )XQF܊LD Nume Prenume $GUHVă WDWăOXL &DVDGH&XOWXUăD 1 MUNICIPIUL VASLUI 1 3UH܈HGLQWH OBREJA NICOLETA I VASLUI, MUNICIPIUL VASLUI Sindicatelor &DVDGH&XOWXUăD 2 MUNICIPIUL VASLUI 1 /RF܊LLWRU APOSTU 0,+$(/$&Ă7Ă/,1$ N VASLUI, MUNICIPIUL VASLUI Sindicatelor &DVDGH&XOWXUăD 3 MUNICIPIUL VASLUI 2 3UH܈HGLQWH LEFTER SIMION C VASLUI, MUNICIPIUL VASLUI Sindicatelor &DVDGH&XOWXUăD 4 MUNICIPIUL VASLUI 2 /RF܊LLWRU &2675Ăù(/ LAURA ù VASLUI, MUNICIPIUL VASLUI Sindicatelor Liceul Teoretic "Mihail 5 MUNICIPIUL VASLUI 3 3UH܈HGLQWH ATASIEI ANDREI T VASLUI, MUNICIPIUL VASLUI .RJăOQLFHDQX Liceul Teoretic "Mihail 6 MUNICIPIUL VASLUI 3 /RF܊LLWRU 52327Ă SIMONA-ANDREEA I VASLUI, MUNICIPIUL VASLUI .RJăOQLFHDQX Liceul Teoretic "Mihail 7 MUNICIPIUL VASLUI 4 3UH܈HGLQWH IVANCIU ILEANA D VASLUI, MUNICIPIUL VASLUI .RJăOQLFHDQX Liceul Teoretic "Mihail 8 MUNICIPIUL VASLUI 4 /RF܊LLWRU 1Ă6758ğ /(18ğ$ M VASLUI, MUNICIPIUL VASLUI .RJăOQLFHDQX Liceul Teoretic "Mihail 9 MUNICIPIUL VASLUI 5 3UH܈HGLQWH IFRIM IOAN I VASLUI, MUNICIPIUL VASLUI .RJăOQLFHDQX Liceul Teoretic "Mihail 10 MUNICIPIUL VASLUI 5 /RF܊LLWRU LAVRIC IONELA-MIRELA I VASLUI, MUNICIPIUL VASLUI .RJăOQLFHDQX 11 MUNICIPIUL VASLUI 6 S.C. COREVAS S.R.L. 3UH܈HGLQWH *5,*25$ù ALEXANDRA D VASLUI, MUNICIPIUL VASLUI 12 MUNICIPIUL VASLUI 6 S.C. COREVAS S.R.L. /RF܊LLWRU PETCULESCU $1'5(($0Ă'Ă/,1$ C VASLUI, MUNICIPIUL VASLUI 13 MUNICIPIUL VASLUI 7 S.C. COREVAS S.R.L. 3UH܈HGLQWH =Ă32'($18 &Ă7Ă/,1 L VASLUI, MUNICIPIUL VASLUI 14 MUNICIPIUL VASLUI 7 S.C.