Emergence Patterns and Latitudinal Adaptations in Development Time of Odonata in North Sweden and Poland
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Week 12 - Day 2
Week 12 - Day 2 Gathering Together by Justin and Amber Paulk For where two or three gather in my name, there I am with them. Matthew 18:20 What a glorious promise from Christ! Come together and He will join you. Whether it’s a happy celebration or a mournful meeting (and certainly all the days in between), how comforting to know that whenever we gather together with others, Christ promises to come along, too. This is what we see in this verse: • God encourages us to gather with others and promises to come alongside our gatherings; • Whenever God is present, so also are His fruits of the spirit including: joy, peace, patience, kindness, goodness, faithfulness, gentleness, self-control, and love First dates can be extremely nerve wracking! “What should I wear?” “Does my hair look good?” “Did I put on deodorant?” “Brush my teeth?” “Is there something sticking out my nose?” All these questions and more fill our minds when we go on first dates—and with good reason—because sometimes our first dates leave a whole lot to be desired! This is one FOTP’s married couple’s account of their awkward first date: “Even though we have been married for 19 years, my wife still makes fun of what I wore on our first date. I had on a green Germany national team soccer jersey with purple jeans. Naturally, they were tight-rolled. (Google it!) In hindsight, I was literally repulsive to look at just based on my color combination! I took her to an Italian restaurant where she chose spaghetti as her main entrée. -

Channel 4 Response to Ofcom's Consultation on 'Protecting Participants in TV and Radio Programmes'
Channel 4 response to Ofcom’s consultation on ‘Protecting participants in TV and radio programmes’ Executive Summary • Channel 4 considers that treating contributors to programmes with due care is of paramount importance, separate to and including our responsibilities under the Ofcom Broadcasting Code (“the Code”).1 • Contributor care has for many years been, and continues to remain, a central priority for us. Working with our production partners we develop comprehensive, proactive and robust welfare protocols and processes which are regularly reviewed and updated. • We take a bespoke approach to contributor care, with the type of support given tailored to the individual nature of each situation. In our experience, what constitutes the proper exercise of our duty of care in each programme depends very much on the nature of the programme, and a ‘one size fits all’ or standard template approach is not appropriate. • As such, we welcome Ofcom’s recognition that “different types of participation may raise very different risks of harm to participants.” And that any proposed new rules “need to be flexible enough to work in a range of situations, and to take account of the fact that very different types and levels of care may be necessary.” • Channel 4 recognises that Ofcom, as the broadcast regulator, has an important role to play in ensuring consistent standards in contributor care across the industry and we believe our current approach is aligned with what Ofcom is seeking to deliver • Whilst we agree that it is vital that Ofcom should be able to provide guidance to the industry and to deal effectively with complaints by viewers and contributors, it must do so within the parameters of the authority granted to it by Parliament and with due regard to other rights and duties including the right to freedom of expression as expressed in Article 10 of the European Convention on Human Rights. -

Observers of First Dates Can Predict Outcome, Study Shows 30 January 2009
Observers of first dates can predict outcome, study shows 30 January 2009 When it comes to assessing the romantic playing brief one-on-one conversations. Each participant field -- who might be interested in whom -- men observed 24 videos, all with different men and and women were shown to be equally good at women, and after each rated whether the man gauging men's interest during an Indiana seemed interested in the woman and the woman in University study involving speed dating -- and the man. equally bad at judging women's interest. The speed dating sessions were all conducted in Researchers expected women to have a leg up in Germany while the observer ratings were all made judging romantic interest, because theoretically by students in Indiana. Despite the language they have more to lose from a bad relationship, but difference, observers were still able to judge men's no such edge was found. romantic interest accurately using body language, tone of voice, eye contact, how often each dater "The hardest-to-read women were being spoke and other non-verbal cues. misperceived at a much higher rate than the hardest-to-read men. Those women were being "How people talk might convey more than what flirtatious, but it turned out they weren't interested they say," Place said. at all," said lead author Skyler Place, a doctoral student in IU's Department of Psychological and Observers did not have to see much of this non- Brain Sciences working with cognitive science verbal behavior. They were just as good at Professor Peter Todd. -

First Dates Sop Casting Rel Final
SLICE™ IS LOOKING FOR SINGLES TO STAR IN NEW CANADIAN DOCUMENTARY SERIES FIRST DATES CANADA Watch a Sneak Peek of First Dates Canada at http://bit.ly/1DwCehs Follow us on Twitter at @shawmediaTV_PR For Immediate Release TORONTO, January 9, 2015 – Force Four Entertainment and Remedy Canada Productions, in association with Shaw Media, are proud to announce the start of production on First Dates Canada, a new Canadian documentary series scheduled to air on Slice in 2015. Based on the hit format from Warner Bros. International Television Production, First Dates Canada brings viewers a compulsively watchable look at the attractions, the heartaches, the connections, and the perils of a first date. Filmed on location in Vancouver, each episode of First Dates Canada documents a restaurant of single men and women who have been paired up for a first date with someone they’ve never met. The restaurant is fitted with more than 40 hidden cameras capturing every moment of the ensuing, and sometimes cringe-worthy, conversational foreplay. Honest and observational at its core, First Dates Canada is a real-life romantic comedy where the players direct themselves. "First Dates Canada is an innovative, entertaining, and incredibly addictive take on the world of modern dating,” said Toby Dormer, President, Remedy Productions Ltd. “We’re sending real singles on real first dates and documenting the drama that unfolds.” "Our production team is already casting Vancouver singles genuinely looking for love,” said John Ritchie, Chief Creative Officer, Force Four Entertainment. “This is a golden opportunity for people of all types, ages, and orientations to have us find their perfect match.” Online applications for First Dates Canada can be filled out by visiting www.Slice.ca/First-Dates-Canada. -

Reading the Irish Woman: Studies in Cultural Encounter and Exchange, 1714–1960
Reading the Irish Woman: Studies in Cultural Encounter and Exchange, 1714–1960 Meaney, Reading the Irish Woman.indd 1 15/07/2013 12:33:33 Reappraisals in Irish History Editors Enda Delaney (University of Edinburgh) Maria Luddy (University of Warwick) Reappraisals in Irish History offers new insights into Irish history, society and culture from 1750. Recognising the many methodologies that make up historical research, the series presents innovative and interdisciplinary work that is conceptual and interpretative, and expands and challenges the common understandings of the Irish past. It showcases new and exciting scholarship on subjects such as the history of gender, power, class, the body, landscape, memory and social and cultural change. It also reflects the diversity of Irish historical writing, since it includes titles that are empirically sophisticated together with conceptually driven synoptic studies. 1. Jonathan Jeffrey Wright, The ‘Natural Leaders’ and their World: Politics, Culture and Society in Belfast, c.1801–1832 Meaney, Reading the Irish Woman.indd 2 15/07/2013 12:33:33 Reading the Irish Woman Studies in Cultural Encounter and Exchange, 1714–1960 GerArdiNE MEANEY, MARY O’Dowd AND BerNAdeTTE WHelAN liVerPool UNIVersiTY Press Meaney, Reading the Irish Woman.indd 3 15/07/2013 12:33:33 reading the irish woman First published 2013 by Liverpool University Press 4 Cambridge Street Liverpool L69 7ZU Copyright © 2013 Gerardine Meaney, Mary O’Dowd and Bernadette Whelan The rights of Gerardine Meaney, Mary O’Dowd and Bernadette Whelan to be identified as the authors of this book have been asserted by them in accordance with the Copyright, Designs and Patents Act 1988. -

Nominations in 2016 Leading Actor Ben Whishaw
NOMINATIONS IN 2016 LEADING ACTOR BEN WHISHAW London Spy – BBC Two IDRIS ELBA Luther – BBC One MARK RYLANCE Wolf Hall – BBC Two STEPHEN GRAHAM This is England ’90 – Channel 4 LEADING ACTRESS CLAIRE FOY Wolf Hall – BBC Two RUTH MADELEY Don’t Take My Baby – BBC Three SHERIDAN SMITH The C-Word – BBC One SURANNE JONES Doctor Foster – BBC One SUPPORTING ACTOR ANTON LESSER Wolf Hall – BBC Two CYRIL NRI Cucumber – Channel 4 IAN MCKELLEN The Dresser – BBC Two TOM COURTENAY Unforgotten - ITV SUPPORTING ACTRESS CHANEL CRESSWELL This is England ’90 – Channel 4 ELEANOR WORTHINGTON-COX The Enfield Haunting LESLEY MANVILLE River – BBC One MICHELLE GOMEZ Doctor Who – BBC One ENTERTAINMENT PERFORMANCE GRAHAM NORTON The Graham Norton Show – BBC One LEIGH FRANCIS Celebrity Juice – ITV2 ROMESH RANGANATHAN Asian Provocateur – BBC Three STEPHEN FRY QI – BBC Two FEMALE PERFORMANCE IN A COMEDY PROGRAMME MICHAELA COEL Chewing Gum – E4 MIRANDA HART Miranda – BBC One SIAN GIBSON Peter Kay’s Car Share – BBC iPlayer SHARON HORGAN Catastrophe – Channel 4 MALE PERFORMANCE IN A COMEDY PROGRAMME HUGH BONNEVILLE W1A – BBC Two JAVONE PRINCE The Javone Prince Show – BBC Two PETER KAY Peter Kay’s Car Share –BBC iPlayer TOBY JONES Detectorists – BBC Four House of Fraser British Academy Television Awards – Nominations Page 1 SINGLE DRAMA THE C-WORD Susan Hogg, Simon Lewis, Nicole Taylor, Tim Kirkby – BBC Drama Production London/BBC One CYBERBULLY Richard Bond, Ben Chanan, David Lobatto, Leah Cooper – Raw TV/Channel 4 DON’T TAKE MY BABY Jack Thorne, Ben Anthony, Pier Wilkie, -

'I Flirt by Insulting People': Exploring the Creation and Performance Of
‘I flirt by insulting people’: Exploring the creation and performance of intimacy in First Dates (Ireland) Brian Clancy Mary Immaculate College, ~University of Limerick~, Ireland First dates Ireland • https://www.youtube.com/watch?v=00PRzwBfIO4 First Dates (Ireland) • 2016-present; • Filmed in Gibson Hotel in Dublin; • Over 20 cameras; • Couples filmed both front and backstage; • At the end of the date, the couples are interviewed together and asked whether they would like to see each other again; • Restaurant closed to the public but you can apply to be a ‘background’ dater; • Each diner given £25 towards the cost of their meal*; • UK, USA, Ireland, Australia, Italy, Canada, Spain, Sweden, Poland, Israel. Dating research • Experimental and reported data (Herz and Inzlicht, 2002; Fisman et al., 2006; Eastwick and Finkel, 2008; Sprecher et al., 2008) • Conversation analysis (del-Teso-Craviotto, 2006; Stokoe, 2010; Korobov, 2011a and b; Korobov, 2017) • Corpus (Korobov, 2011a and b; Korobov, 2017) The dates Name S/U Words Introduction (narrator) Amy (21) & S 2242 Legal student Amy has tried a few partners to date James (25) none have been judged worthy. Simon (36) & S 1276 - Tynan (39) Federica (25) & U 1825 South African model Federica likes to take the lead Lee (33) her dance partner will need stamina. Mark (39)& U 1496 Like any seasoned warrior Mark appreciates the Gemma (39) value of a positive mental attitude. Mark's blind date is clued in single mum Gemma. Dawn (24) & U 1429 Free spirited Dawn is a real girl's girl but she's not Alannah (21) just any girl's girl. -
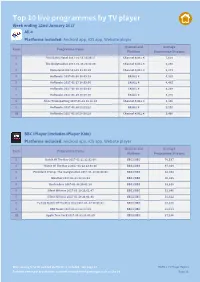
Top 10 Live Programmes by TV Player Week Ending 22Nd January 2017 All 4 Platforms Included: Android App, Ios App, Website Player
Top 10 live programmes by TV player Week ending 22nd January 2017 All 4 Platforms included: Android app, iOS app, Website player Channel and Average Rank Programme Name Platform Programme Streams 1 First Dates Hotel 2017-01-16 22:03:17 Channel 4/ALL 4 7,628 2 The Undateables 2017-01-16 21:02:00 Channel 4/ALL 4 6,298 3 Homeland 2017-01-22 21:02:25 Channel 4/ALL 4 5,273 4 Hollyoaks 2017-01-16 18:59:29 E4/ALL 4 4,915 5 Hollyoaks 2017-01-17 18:59:06 E4/ALL 4 4,465 6 Hollyoaks 2017-01-18 18:59:03 E4/ALL 4 4,299 7 Hollyoaks 2017-01-19 18:59:10 E4/ALL 4 4,276 8 Film: Trainspotting 2017-01-22 22:06:09 Channel 4/ALL 4 3,995 9 Hollyoaks 2017-01-20 18:59:10 E4/ALL 4 3,554 10 Hollyoaks 2017-01-16 18:30:28 Channel 4/ALL 4 3,488 BBC iPlayer (includes iPlayer Kids) Platforms included: Android app, iOS app, Website player Channel and Average Rank Programme Name Platform Programme Streams 1 Match Of The Day 2017-01-21 22:32:30 BBC1/BBC 70,537 2 Match Of The Day 2 2017-01-22 22:30:16 BBC1/BBC 47,014 3 President Trump: The Inauguration 2017-01-20 16:00:00 BBC1/BBC 42,084 4 Weather 2017-01-21 22:28:43 BBC1/BBC 40,035 5 Eastenders 2017-01-16 20:01:14 BBC1/BBC 33,108 6 Silent Witness 2017-01-18 21:01:47 BBC1/BBC 32,940 7 Silent Witness 2017-01-16 21:01:35 BBC1/BBC 30,822 8 Fa Cup Match Of The Day Live 2017-01-17 19:59:00 BBC1/BBC 28,821 9 BBC News 2017-01-21 22:12:15 BBC1/BBC 28,414 10 Apple Tree Yard 2017-01-22 21:01:25 BBC1/BBC 27,286 Only viewing time for audited platforms is included. -

Best Italian Restaurants in Chicago"
"Best Italian Restaurants in Chicago" Created by: Cityseeker 12 Locations Bookmarked Caffe Baci "Tasty Cafe and Top Caterer" Enjoy Caffe Baci at either one of its five locations or for a catered event. The Cafe is established as a top Chicago caterer for corporate meetings and special events, but is also a great sit-down cafe for breakfast and lunch. Choose from a selection of pastas, chicken entrees, pizza, soup and sandwiches such as the Curried Couscous, Spinach Ravioli, Campanella with Diced Chicken & Artichoke, Shrimp Bisque or the Mozza e Pom by Caglar Araz on Unsplash Panini sandwich. Check out the website for daily specials at each location. +1 312 629 2215 www.caffebaci.com [email protected] 2 North LaSalle Street, Chicago IL Coco Pazzo "Italian Feast" Many people in town, food critics and laymen alike, rank this as the best Italian restaurant in the city. Coco Pazzo has a wonderful menu, mixing a variety of styles and flavors to create Tuscan inspired dishes. The menu changes often to ensure the food is only prepared with the freshest ingredients. Pair you meal with a glass of wine from their impressive by bady qb on Unsplash collection of over 150 wines. Make your reservation well in advance and dress to impress. +1 312 836 0900 www.cocopazzochicago.c jweiss@cocopazzochicago. 300 West Hubbard Street, om com Chicago IL Rosebud on Rush "Local Talent" Famed local Chicagoan Alex Dana, founder of the Rosebud Restaurant Group has been serving diners in the Windy City since 1973. With the exception of Mama's Boy and Carmine's, most of his eateries around the Chicagoland area carry the Rosebud name, but one thing is for certain, all of them provide the same excellent service and massive portions on every plate. -

Study on the First Arrival Date of Spring Avian Migrants to Eastern Poland
Polish J. of Environ. Stud. Vol. 20, No. 4 (2011), 843-849 Original Research Study on the First Arrival Date of Spring Avian Migrants to Eastern Poland Waldemar Biaduń1*, Ignacy Kitowski2**, Eugeniusz Filipiuk3 1Department of Zoology and Invertebrate Ecology, Catholic University of Lublin, Al. Kraśnicka 102, 20-718 Lublin, Poland 2Department of Zoology, University of Life Sciences in Lublin, Akademicka 13, 20-950 Lublin, Poland 3Department of Meteorology and Climatology, Institute of Earth Sciences, Maria Curie-Skłodowska University, Al. Kraśnicka 2cd, 20-718 Lublin, Poland Received: 9 July 2010 Accepted: 23 February 2011 Abstract First arrival dates of birds to their breeding grounds can be a valuable biological trait reflecting global and local climate changes. Long-term trends in the first arrival dates of 28 common breeding bird species to Gułowskie Forests (eastern Poland) were studied for the period of 1970-2009. Only 6 species studied – Barn Swallow (18 days), Thrush Nightingale (28 days), Lesser Whitethroat (8 days), Golden Oriole (7 days), Common Blackbird (13 days), and Chaffinch (14 days) – demonstrated a statistically significant trend of ear- lier arrival. Short- and medium-distance migrants arrived significantly earlier (median date: March 22) than studied long-distance migrants (median: April 23). Keywords: migration, phenology, first arrival dates, migrants, Gułowskie Forests, Poland Introduction The aim of this study is to analyze trends in the first arrival date (FAD) of 28 common breeding avian species to Numerous species of vertebrates have advanced their Gułowskie Forests (eastern Poland) from 1970 to 2009. phenology in recent decades. Many authors indicate that These analyses were performed in the context of climatic these processes are related to climate change [1-6]. -

Read Book the Swedish Girl
THE SWEDISH GIRL: 10 PDF, EPUB, EBOOK Alex Gray | 480 pages | 12 Dec 2013 | Little, Brown Book Group | 9780751548228 | English | London, United Kingdom The Swedish Girl: 10 PDF Book The plot thickens as it is discovered that the victim, the Swedish girl, was very promiscuous and had sex with another roommate, Colin, prior to her murder. Votes: 12, Filming was projected to start in Spring in Berlin. Too bad it was all a facade. Deadline Hollywood. School strike for climate. Archived from the original on 5 October Archived from the original on 31 October It doesn't take long for a new murder investigation only this time he has to wait for his team to keep him updated. Archived from the original on 27 September European Parliament Press release. Choose an adventure below and discover your next favorite movie or TV show. Royal Scottish Geographical Society. Best Asian Brides. Websites like Culture Trip or even looking through the suggestions on Google Maps are your best friend in this. This is probably one of the best books I have ever read. Archived from the original on 27 January We scheduled various directors and with each director came a new draft. Over and over again. The Danish Girl by David Ebershoff. However a young girl is murdered in the house she lives in. Archived from the original on 3 October In this article, we will attempt to elucidate the confusion related to the most famous Scandinavian women and shed light on how to be successful with dating Swedish girls. Many girls are obsessed with American culture, lifestyle, and the look of the US guys. -

Important Fitness Al., 2000). Time of Is Impor
Odonatologica39(2): 97-106 June 1, 2010 Emergence patterns and latitudinaladaptations in developmenttime of Odonata in North Sweden and Poland F. Johansson¹,S. Śniegula²and T. Brodin¹ 1 Department of Ecology and Environmental Science, Umefi University, SE-90187 Umefi, Sweden 2 Department of Ecosystem Conservation, Institute of Nature Conservation, Polish Academy of Sciences, Mickiewicza 33, PO-31-120 Krakow, Poland Received December 17, 2009 / Revised and Accepted January 26, 2010 data dates of from north- Using exuviae, are presented on emergence dragonflies Sweden and northwestern Poland. The 17 in Sweden showed ern spp. sampled con- siderable overlap in emergence periods. In Sweden, Leucorrhinia rubicunda was the A first sp. to emerge (May 31) and Sympetrum danae the last (July 19). comparisonof first dates of of in Sweden Poland showed difference emergence spp. and a between 9 and 30 with all Polish first. days, spp. emerging Compared tospring species, summer species and obligate univoltine summer species showed less difference in first date of between Swedish and Polish emergence populations. In a laboratoryexperimentLeu- corrhinia dubia reared from both from the final instar larva under was regions egg to northern Swedish and northwestern Polish photoperiods. Swedish larvae developed fasterunder anorthern Swedish photoperiodcomparedto anorthwestern Polish pho- toperiod. However, no such difference in developmentwas found for northwestern This differences Polish larvae. suggests that there aregenetic between both populations in response to photoperiod.The results are discussed in the context of compensation of larval developmentof northern populationsin relation to photoperiod. INTRODUCTION Time of and larval size at fitness emergence emergenceare important compo- nents in dragonflies (ANHOLT, 1991; DE BLOCK & STOKS, 2005; BANKS & THOMPSON, 1987; SOKOLOVSKA et al., Timeof is 2000).