Leatherbacks Gabon Stephen K. Pikesley A,B, Pierre Didier
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

12. Monte Alén-Monts De Cristal Landscape
12. Monte Alén-Monts de Cristal Landscape Figure 12.1. Map of Monte Alén-Monts de Cristal Landscape (Sources: CARPE, JRC, SRTM, WCS-Gabon). Location and area he Monte Alén-Monts de Cristal Landscape Th e Landscape in brief Tcovers the south and southeast of Equatorial Guinea and the northwest of Gabon (Figure 12.1). Coordinates: 1°53’35’’N – 0°5’38’’N; 9°37’2’’E – 11°36’3’’E It has an area of approximately 26,747 km2, of Area: 26,747 km2 which about half is located in Equatorial Guinea Elevation: 300-1,250 m and half in Gabon. In Equatorial Guinea, it in- Terrestrial ecoregion: Atlantic Congolese forests ecoregion cludes the Monte Alén and Altos de Nsork na- Aquatic ecoregions: Central West equatorial coastal ecoregion tional parks, as well as the Rio Muni Estuary Southwest equatorial coastal ecoregion Reserve and the Piedra Nzas Natural Monument. Protected areas: In Gabon, it comprises the two sections of Monts Monte Alén National Park, 200,000 ha, 1988/2000, Equatorial Guinea de Cristal National Park. Altos de Nsork National Park, 40,000 ha, 2000, Equatorial Guinea Monts de Cristal National Park, 120,000 ha, 2002, Gabon Physical environment Rio Muni Estuary Reserve, 70,000 ha, 2000, Equatorial Guinea Piedra Nzas Natural Monument, 19,000 ha, 2000, Equatorial Guinea Relief and altitude Th e Landscape occupies a rugged area of pla- teaus and mountain chains mainly situated at an altitude of 300 m to 650 m to the northeast of the coastal sedimentary basin of Gabon (Figure 12.2). In Equatorial Guinea, the highest peak is formed by Monte Mitra, which rises to 1,250 m and is 114 the culminating point of the Niefang chain which runs from the southwest to the northeast. -
1. Introduction 86
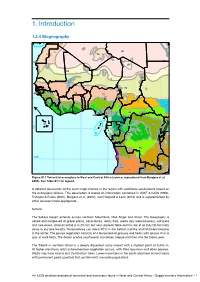
34 85 1. Introduction 86 69 30N 60 1.2.4 Biogeography87 88 93 65 95 98 96 92 97 94 111 99 62 61 35 100 101 115 36 25 70 2 39 83 102 37 38 59 71 1 4 3 4 6 7 5 10 40 44 116 9 103 104 31 12 11 13 16 0 73 41 8 18 14 45 15 17 66 20 47 72 27 43 48 46 42 19 118 112 10S 81 74 50 21 52 82 49 32 26 56 Figure S1.1 Terrestrial ecoregions in West and Central Africa (source: reproduced from Burgess et al. 75 2004). See Table S1.1 for legend. 106 51 119 33 55 64 53 67 63 84 A2 0detailedS description of the eight major biomes in the region with additional subdivisions based on76 29 the ecoregions follows. This description is based on information contained 5in8 WWF6 8& IUCN (1994), 30 Fishpool & Evans (2001), Burgess et al. (2004), and Happold & Lock (2013) and is supplemented57 by 114 other sources where appropriate. 107 54 Terrestrial ecoregions 105 109 113 Sahara Country boundary 22 77 78 The Sahara Desert extends across northern Mauritania, Mali, Niger and Chad. The topography11 7is 28 30S 79 varied and composed of gravel plains, sand dunes, rocky flats, wadis110 (dry watercourses), salt pans 108 23 and rare oases. Annual rainfall is 0–25 mm but very unpredictable and no rain at all may fall for many 0 250 500 1,000 1,500 2,000 2,500 80 years in any one locality. -

Ivindo National Park: Resource Use on the Ivindo River Corridor
Ivindo National Park: Resource Use on the Ivindo River Corridor Progress Report and Draft Strategy Document Matthew Steil Ivindo National Park Wildlife Conservation Society Acknowledgements This resource use study and related implementation activities are made possible through financial support from the United States Agency for International Development (USAID) through its Central African Regional Program for the Environment (CARPE). All viewpoints expressed within are uniquely those of the author and do not necessarily reflect those of USAID, Gabon’s National Park Agency or the Wildlife Conservation Society. 2 Preface This document is a progress report prepared for USAID-CARPE on ongoing activities to implement a participatory resource use management plan for fishing communities using the Ivindo River corridor passing through the northern section of Ivindo National Park. Work is continuing through FY08 and following completion of field activities, a more comprehensive set of recommendations for the use of this corridor based on findings from the study and participatory stakeholder meetings will be detailed. 3 Background General The Ivindo River is one of two major tributaries of the Ogooué and is responsible for draining most of northeastern Gabon, along with areas of northern Congo and Southern Cameroon. Due in large part to the significant organic decomposition in the waters of its tributaries, Ivindo is one of the only large blackwater rivers in the sub-region. Currently, the Ivindo River Basin drains what is either primarily intact forest or sparsely populated areas, with the Gabonese provincial capital Makokou (pop ~ 15,000) as the only major populated area within the basin. The Ivindo River Basin covers 62,700 km2 and drains a significant area of the TRIDOM landscape, containing within it parts of three National Parks in Gabon: Minkebé, Mwagna and Ivindo (Map 1). -

Our Vision for Conservation in Central Africa
U.S. Fish & Wildlife Service A Results-Based Vision for Conservation in Central Africa Acknowledgments Contributions to the development of this document were provided by: Bryan Arroyo, Gloria Bell, Dirck Byler, Daphne Carlson Bremer, Michelle Gadd, Nancy Gelman, Tatiana Hendrix, Danielle Kessler, Frank Kohn, Earl Possardt, Herb Raffaele, Krishna Roy, Richard Ruggiero and Jim Serfis of the U.S. Fish and Wildlife Service (USFWS). External reviews were made by: Jason Ko, U.S. Forest Service; Sanath K. Reddy, U.S. Agency for International Development / Central Africa Regional Program for the Environment (USAID / CARPE); Tim Resch, USAID Africa Bureau; and Matthew Cassetta, U.S. Department of State. Edited by: William Kanapaux, Matthew Muir and Heidi Ruffler Layout and design by: Heidi Ruffler, Mark Newcastle and Tatiana Hendrix International Affairs U.S. Fish and Wildlife Service MS: IA 5275 Leesburg Pike Falls Church, VA 22041-3803 www.fws.gov/international Front and back cover: Mountain gorilla family group in Central Africa. Credit: Dirck Byler / USFWS 2 A Vision for Central Africa A Vision for Central Africa 3 4 A Vision for Central Africa Table of Contents Wildlife Areas in Central Africa with USFWS Investments, 2011-2014................................6 A Vision for Central Africa............................................................................................................7 A Strategic Framework for Central Africa.................................................................................8 From exploitation to conservation: -

Preliminary Assessment of the Frog Assemblages from Sites Adjacent to Three National Parks in Gabon
Herpetological Conservation and Biology 13(1):240–256. Submitted: 8 August 2017; Accepted: 1 March 2018; Published 30 April 2018. PRELIMINARY ASSESSMENT OF THE FROG ASSEMBLAGES FROM SITES ADJACENT TO THREE NATIONAL PARKS IN GABON JOANNA G. LARSON1,2,3 AND BREDA M. ZIMKUS2 1Department of Ecology and Evolutionary Biology and Museum of Zoology, University of Michigan, Ann Arbor, Michigan 48109, USA 2Museum of Comparative Zoology, Harvard University, Cambridge, Massachusetts 02138, USA 3Corresponding author, e-mail: [email protected] Abstract.—We report on preliminary frog inventories from three sites in Gabon that are located in close proximity to three national parks. In the lowland forest surrounding Ossélé Village, located north of Batéké Plateau National Park, we documented 14 species from nine genera and six families. The species assemblage within the area of Ossélé Village was markedly different from what is known in the Batéké Plateau National Park from recent inventory work, likely due to the secondary rainforest located outside of the national park and the savanna grassland interspersed with gallery forest within the protected area. We recorded 10 species from eight genera and five families in the buffer zone of Birougou National Park. From the buffer zone of Minkébé National Park, we documented 16 species from eight genera and six families. The majority of these species are widely distributed in the lowland forests of Central Africa. No amphibian surveys have yet been undertaken within Birougou and Minkébe National Parks, but the information provided by these inventories provides insight into species that are likely present since the habitat in these buffer zones is found in the adjacent protected areas. -

Amphibians and National Parks in Gabon, Western Central Africa Amphibien Und Nationalparks in Gabun, Westliches Zentralafrika
©Österreichische Gesellschaft für Herpetologie e.V., Wien, Austria, download unter www.biologiezentrum.at HERPETOZOA 19 (3/4): 135 - 148 135 Wien, 30. Jänner 2007 Amphibians and National Parks in Gabon, western Central Africa Amphibien und Nationalparks in Gabun, westliches Zentralafrika OLIVIER S. G. PAUWELS & MARK-OLIVER RODEL KURZFASSUNG Wir geben eine Zusammenfassung des derzeitigen Wissens über die Amphibieninventare der in Gabun in jüngster Zeit eingerichteten Nationalparks. Vorläufige Arteninventare sind nur für fünf der 13 Parks vorhanden: Crystal, Ivindo, Loango, Lopé und Moukalaba-Doudou. Sechsundsiebzig (86%) der 88 aus Gabun bekannten Arten, alle 10 (100%) der fast endemischen, und drei der sechs (50%) für Gabun endemischen Arten sind aus diesen Nationalparks nachgewiesen. Vorrangig sollte es sein, die Verbreitung der Amphibien in Gabun durch intensive Surveyarbeit besser zu verstehen und neue Schutzgebiete für die Arten zu schaffen, deren Vorkommen noch nicht ausreichend durch die bestehenden Nationalparks abgedeckt sind. ABSTRACT A synthesis of the current state-of-knowledge of amphibian diversity in Gabon and in the recently created Gabonese National Parks is provided. Preliminary inventories are available for five of the 13 parks: Crystal, Ivindo, Loango, Lopé and Moukalaba-Doudou. Seventy-six (86%) of the 88 species known to occur in Gabon, all ten near-endemics (100%), and three of the six Gabonese endemic species (50%) are currently represented in these parks. Future priority actions should comprise an intensified survey -

MARINE TURTLES in ATLANTIC AFRICA Editorials: Proceeding to the Future
Issue Number 116 April 2007 Measuring a male green turtle captured by Benga fishermen in Gabon (photo by Alejandro Fallabrino). SPECIAL FOCUS ISSUE - MARINE TURTLES IN ATLANTIC AFRICA Editorials: Proceeding to the Future..................................................................................................................................M.S. Coyne The Oldest Place Where There is Always Something New............................................................................J.G. Frazier Articles: An Update on Marine Turtles in Côte d’Ivoire, West Africa .................................................................J.G. Peñate et al. An Update on Sea Turtle Conservation Activities in the Republic of Congo..................................................G. Bal ������et al. Conflicts & Social Dilemmas Associated with Capture of Marine Turtles by Artisanal Fishers in Benin.....J.S. Dossa et al. Marine Turtle Mortality in Southern Gabon and Northern Congo............................................................R. Parnell et al. Artificial Lights as a Significant Cause of Morbidity of Leatherbacks in Pongara Natl. Park, Gabon.....S.L. Deem et al. Observations of Marine Turtles in Relation to Seismic Airgun Sound off Angola............................................C.R. Weir Fibropapillomatosis confirmed in Chelonia mydas in the Gulf of Guinea,West Africa...........................A. Formia et al. Presence of Sea Turtles in Traditional Pharmacopoeia and Beliefs of West Africa....................................J. Fretey et al. -

Review of Progress on the Regional Action Plan for the Conservation of Chimpanzees and Gorillas in Western Equatorial Africa
UNEP/GA/MOP2/Inf.1.4 Agenda Item 6.0 Review of progress on the Regional Action plan for the Conservation of Chimpanzees and Gorillas in Gabon Compiled by Fiona Maisels, 2010 Updated by Natacha Ona and Kathryn Jeffery, 2011 Contributors: Gaspard Abitsi, Kathryn Jeffery, Fiona Maisels, Tim Rayden, Richard Parnell, Sandra Ratiarison, Patricia Reed, Aimee Sanders, Malcolm Starkey, Ruth Starkey, Richard Zanre Overview ................................................................................................................. 1 What has been done since 2005? ......................................................................... 6 Exceptional Priority Areas .................................................................................................. 7 1. Loango/ Moukalaba-Doudou/ Gamba complex, Gabon ....................................................................... 7 2. Lopé/ Waka complex, Gabon ...............................................................................................................10 Important Priority Areas ................................................................................................... 13 3. Ivindo complex, Gabon ........................................................................................................................13 4. Belinga-Djoua, Gabon .........................................................................................................................16 5.Conkouati/Mayumba complex, Republics of Congo, Gabon ................................................................17 -

Marine Turtle Update
MARINE TURTLE UPDATE Recent News from the WWF Africa & Madagascar Marine Turtle Programme © WWF-Canon / Jürgen FREUND Number 2 – November 2005 In 2002 WWF launched a new Africa and Madagascar Marine Turtles Programme. Building on over 30 years of experience in marine turtle conservation, WWF’s new initiative aims to provide strategic field interventions to help guarantee a future for these threatened species. The long term goal of the programme (25 years) is: Viable populations of all five species of Marine Turtles in African waters are conserved. The Programme has four targets 1. Loss and degradation of critical nesting, inter-nesting and foraging habitats reduced or prevented in at least eight key sites by 2010. 2. Measures to control unsustainable use and trade of marine turtles and turtle products enhanced in at least six countries by 2010. 3. Incidental capture of marine turtles reduced in the territorial waters of at least six countries and in at least two pelagic fisheries by 2010. 4. Capacity for monitoring, research and management of marine turtles and their habitats enhanced in at least eight countries by 2006. For further information on the WWF Africa and Madagascar Marine Turtles Programme, check our website: http://www.panda.org/africa/marineturtles or contact: Sarah Humphrey Africa and Madagascar Programme WWF International Avenue du Mont Blanc CH 1196 Gland Switzerland Tel: +41 22 364 9111 Email: [email protected] This edition of Marine Turtle Update was compiled and edited by Sarah Humphrey & Alison Wilson. Layout by Sandrine Jiménez. Marine Turtle Update provides recent news on the conservation work undertaken and supported by WWF in Africa and Madagascar to conserve marine turtles. -

Wildlife Monitoring Practices and Use in Central Africa
Program on African Protected Areas & Conservation (PAPACO) Wildlife monitoring practices and use in Central Africa By Starkey, M., Scholtz, O. and Taylor, G. Wildlife Conservation Society Executive Summary Monitoring is an essential part of adaptive management, and is necessary for evaluating the outcomes of conservation action. Ecological monitoring plays other roles in conservation, serving as an important communication tool for illustrating the plight of a species, demonstrating the outcomes of payment for ecosystem services schemes, and testing the success of different types of protected areas. Conversely monitoring is not a panacea; good ecological data in itself does not ensure the protection of a species. Central Africa harbours an amazing diversity in both forest and savanna ecosystems. Over 153 protected areas have been created, covering more than 423,000 km2, with ecological monitoring an increasingly important theme at the regional level. In the context of the increasing recognition of the value and plight of Central Africa’s protected areas and the role of ecological monitoring, this project conducted a comprehensive collation of wildlife monitoring reports from Central Africa’s protected areas; the goal was to evaluate the status and uses of wildlife monitoring, and identify the challenges and opportunities for improving this. A database of protected areas and their ecological monitoring information has been completed, with preliminary analyses presented in this report. Our final sample included 121 protected areas (PAs) in six countries in Central Africa, with 205 reports on 255 individual surveys meeting the monitoring criteria we had set. We did not receive any response (“no information”) from 33 PAs, with most of these in the Democratic Republic of Congo. -

13. Gamba-Mayumba-Conkouati Landscape
13. Gamba-Mayumba-Conkouati Landscape Figure 13.1. Map of Gamba-Mayumba-Conkouati Landscape (Sources: CARPE, JRC, SRTM, WCS-Gabon). Location and area Th e Landscape in brief he Gamba-Mayumba-Conkouati Landscape Tis transnational and centered on the Loango, Coordinates: 1°36’26’’S – 4°26’26’’S; 9°15’48’’E – 12°24’28’’E Moukalaba-Doudou and Mayumba national Area: 34,258 km2 parks in Gabon, and Conkouati-Douli National Elevation: 0-840 m Park in the Republic of Congo (Figure 13.1). Land ecoregions: Lying between the national parks in Gabon is a Congolese Atlantic forests ecoregion set of hunting areas - Ngové-Ndogo, Moukalaba, Southwestern forest-savannah mosaic ecoregion Sette Cama and Iguéla - plus the Ouanga Plain Aquatic ecoregion: Wildlife Reserve. Th e Landscape extends over a Southernmost western equatorial coastal ecoregion total area of 34,258 km², of which approximately Protected areas: 75% lies in Gabon and 25% lies in the Republic Loango National Park *, 153,581 hectares, 2002, Gabon of Congo, and stretches along the southwestern Moukalaba-Doudou National Park *, 502,805 hectares, 2002, Gabon coast of Gabon and the western coast of the Mayumba National Park, 80,000 hectares, 2002, Gabon Republic of Congo. Mayumba and Conkouati- Conkouati-Douli National Park, 505,000 hectares, 1980/1999, Republic of Douli national parks extend back 15 km and Congo 22 km from the beach respectively, covering an Ngové-Ndogo Hunting Area*, 1956, Gabon area of 80,000 hectares in Gabon and 120,000 Moukalaba Hunting Area *, 20,000 hectares, 1962, Gabon hectares in the Republic of Congo. -

Four Years of Marine Turtle Monitoring in the Gamba Complex Of
Four Years of Marine Turtle Monitoring in the Gamba Complex of Protected Areas Gabon, Central Africa 2002-2006 Bas Verhage, Eustache Beodo Moundjim, Suzanne Rachel Livingstone Financial support from: In cooperation with: and: PROTOMAC Four years of Marine Turtle Monitoring in the Gamba Complex, Gabon Table of contents Acronyms...................................................................................................................... 3 Abstract......................................................................................................................... 4 1 Introduction ............................................................................................................... 5 1.1 General introduction ......................................................................................... 5 1.2 Stakeholder mapping........................................................................................ 6 1.3 Objectives of the project................................................................................... 7 1.4 Sea turtles in Western/Central Africa .............................................................. 8 1.5 Protection........................................................................................................... 9 1.6 Research area - The Gamba Complex ............................................................. 9 1.7 Study site ......................................................................................................... 10 2 Project activities ....................................................................................................